
 French
French Deutsch
DeutschHomo erectus – Wikipedia
| Homo erectus | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
 Homo-erectus-Skelett des etwa neun Jahre alten | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Pleistozän | ||||||||||||
| 2,0 bis ? Mio. Jahre | ||||||||||||
| Fundorte | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Homo erectus | ||||||||||||
| (Dubois, 1893) | ||||||||||||
Homo erectus ist eine ausgestorbene Art der Menschenaffen-Gattung Homo. Aus den Homo erectus zugeschriebenen pleistozänen Populationen Afrikas entwickelte sich vermutlich in Europa der Neandertaler und – parallel zu diesem, aber unabhängig von ihm – in Afrika der anatomisch moderne Mensch (Homo sapiens). Laut Richard Leakey war Homo erectus „die erste hominine Art, die das Feuer benutzte; die erste, die das Jagen als ein wesentliches Element zur Sicherung ihrer Nahrungsversorgung einsetzte; die erste, die wie ein moderner Mensch laufen konnte.“[1] Die Abgrenzung des Homo erectus von anderen Arten der Gattung Homo ist in Fachkreisen – zwischen sogenannten Lumpern und Splittern – allerdings umstritten. Zudem entwickelten sich aus Homo erectus – unabhängig voneinander – sowohl Homo heidelbergensis und Neandertaler (in Eurasien) als auch Homo sapiens (in Afrika) und vermutlich Homo floresiensis (in Ostasien). Ein „Aussterbezeitpunkt“ ist daher nicht datierbar (vergleiche die Grafiken weiter unten).
Die ersten Fossilien des Homo erectus wurden ab den 1890er-Jahren in Asien entdeckt. Dies führte jahrzehntelang dazu, dass die Paläoanthropologen zu dem Schluss kamen, der anatomisch moderne Mensch habe sich in Asien aus affenähnlichen Vorfahren entwickelt, obwohl bereits Charles Darwin 1871 vermutet hatte,[2] der Mensch habe sich in Afrika entwickelt, da seine nächsten Verwandten – Schimpansen und Gorillas – dort beheimatet sind. Diese Einschätzung verhinderte zudem zwei Jahrzehnte lang die Anerkennung des ersten afrikanischen Australopithecus-africanus-Fundes – des 1924 geborgenen Kindes von Taung – als zugehörig zur Ahnenreihe des modernen Menschen. Als wahrscheinlicher Irrtum wurde dies erst erkannt, nachdem in Afrika entdeckte, sehr alte hominine Fossilien im April 1964 von Louis Leakey als Homo habilis an die Basis der Gattung Homo gestellt worden waren.[3]
Namensgebung
Die Bezeichnung der Gattung Homo ist von lateinisch homo [], deutsch ‚Mensch‘ abgeleitet. Der Artname erectus ist von lateinisch erigere („aufrichten“) abgeleitet. Homo erectus bedeutet somit „der aufgerichtete Mensch“.
Die ersten drei Fossilien, Trinil I bis III, die 1891/92 von Eugène Dubois entdeckt und als Java-Mensch bekannt wurden, hatte Dubois zunächst als Anthropopithecus erectus („aufgerichteter Mensch-Affe“) benannt, abgeleitet von altgriechisch ἄνθρωπος anthropos, deutsch ‚Mensch‘ und griech. πίθηκος, altgr. ausgesprochen píthēkos („Affe“).[4] 1894 änderte Dubois den Gattungsnamen in einem Fachaufsatz in Pithecanthropus erectus („aufgerichteter Affen-Mensch“)[5] und beschrieb die Art als Übergangsform, die zum modernen Menschen geführt habe. Dubois griff bei dieser Namensgebung einen Vorschlag auf, den Ernst Haeckel 1863 empfohlen hatte, als er eine hypothetische Übergangsform zwischen Mensch und Affe beschrieben und diese Pithecanthropus genannt hatte.[6]
1944 publizierte Theodosius Dobzhansky den Aufsatz On species and races of living and fossil man, in dem er argumentierte, dass es zu jeder Zeit nur eine einzige, variationsreiche Art der Hominini gegeben habe.[7] 1950 unterstützte Ernst Mayr in einem Vortrag beim Cold Spring Harbour Symposium on Quantitative Biology Dobzhanskys Annahmen und überzeugte die Paläoanthropologen davon, alle inzwischen entdeckten, mutmaßlichen Vorfahren des Menschen der Gattung Homo zuzuordnen und künftig darauf zu verzichten, Einzelfunde mit einem Gattungs- und Artnamen zu benennen; stattdessen solle man solche Fossilien nach ihrer Herkunft benennen (zum Beispiel Sterkfontein-Fund statt „Plesianthropus“ für Mrs. Ples).[8] Dies hatte zur Folge, dass auch die als Pithecanthropus erectus bezeichneten Fossilien erneut umbenannt wurden, dass gemäß den internationalen Regeln für die Zoologische Nomenklatur der Artname erectus erhalten blieb und die von Eugène Dubois entdeckten drei Fossilien nunmehr die Typusexemplare von Homo erectus sind.
Erstbeschreibung
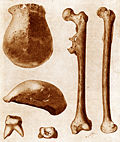
Als Holotypus von Homo erectus wurden 1893 drei von Eugène Dubois auf der indonesischen Insel Java geborgene Fossilien benannt: ein im Oktober 1891 entdecktes Schädeldach mit einem markanten, durchgehenden Querwulst des Stirnbeins oberhalb der Nasenwurzel („Überaugenwulst“), Sammlungsnummer Trinil II; ein im August 1892 entdeckter, vollständig erhaltener Oberschenkelknochen (Trinil III), der in Form und Aufbau dem Oberschenkelknochen des anatomisch modernen Menschen gleicht, was von Dubois als Hinweis gedeutet wurde, dass sein Besitzer sich ähnlich aufrecht wie der moderne Mensch bewegte; sowie ein bereits im August 1891 entdeckter großer Backenzahn (Trinil I).
Zahn und Schädeldach waren von Dubois zunächst als Fossil eines nicht-homininen Menschenaffen interpretiert worden, weil Dubois eine Ähnlichkeit mit den rezenten Schimpansen zu erkennen glaubte. Daher ordnete er seine Funde zunächst der neuen Art Anthropopithecus erectus zu; Anthropopithecus war damals der Gattungsname der Schimpansen.
Die von Eugène Dubois entdeckten drei Fossilien galten damals als die ersten Funde von Fossilien der Hominini außerhalb Europas und zugleich als die ältesten bis dahin aus dieser Klade entdeckten. Aufgrund ihrer Fundsituation am Rande eines ehemaligen Gewässers ist die Datierung der Knochen problematisch; nach heutigem Kenntnisstand wird dem Schädeldach ein Alter von rund einer Million Jahre zugeschrieben. Allerdings besitzt das Schädeldach so wenige charakteristische Merkmale, dass man es nicht zuverlässig mit anderen Fossilien vergleichen kann.[9] Die Zuordnung des Backenzahns zur Gattung Homo gilt heute hingegen als nicht gesichert,[10] und der Oberschenkelknochen – dessen Gestalt ursächlich war für die Wahl des Artnamens erectus – wird heute als vermutlich dem anatomisch modernen Menschen (Homo sapiens) zugehörig interpretiert:[11] Eine Deutung, die der britische Anatom Daniel John Cunningham (1850–1909) bereits 1895 im Fachblatt Nature publiziert hat.[12]
Die Erstbeschreibung in Dubois’ Veröffentlichung Paleontologische onderzoekingen op Java ist im Original datiert auf das Jahr 1892, weswegen die vollständige Bezeichnung der Art lange Zeit Homo erectus DUBOIS, 1892 lautete. Bert Theunissen wies allerdings 1989 darauf hin, dass die Veröffentlichung erst 1893 erschienen sei, daher müsse die Art Homo erectus DUBOIS, 1893 benannt werden.[13][14] 2024 wurde dieser Hinweis von einer anderen Autorengruppe in einem Review-Artikel bestätigt;[15] zugleich wurde bekräftigt, dass wegen der unsicheren Zuordnung der Fossilien Trinil I und III nur das Schädeldach Trinil II als Holotypus infrage kommt.
-
-
 Schädeldach „Sangiran II“, Original. Sammlung Koenigswald im Naturmuseum Senckenberg.
Schädeldach „Sangiran II“, Original. Sammlung Koenigswald im Naturmuseum Senckenberg.
Man beachte den Überaugenwulst über dem linken Auge. -
 Sangiran IV: Oberkiefer mit Gaumen (Original, 1,6 mya; Sammlung Koenigswald im Naturmuseum Senckenberg)
Sangiran IV: Oberkiefer mit Gaumen (Original, 1,6 mya; Sammlung Koenigswald im Naturmuseum Senckenberg) -
 Sangiran 17 = Pithecanthropus VIII = Homo erectus, entdeckt 1969 (Kopie)
Sangiran 17 = Pithecanthropus VIII = Homo erectus, entdeckt 1969 (Kopie) -
 Unterkiefer von Ternifine, zunächst bezeichnet als Atlanthropus mauritanicus
Unterkiefer von Ternifine, zunächst bezeichnet als Atlanthropus mauritanicus

.jpg)
.jpg)


Alter
Homo erectus werden Fossilien aus dem Pleistozän zugeordnet, deren älteste rund zwei Millionen Jahre alt sind und somit dem frühesten Altpleistozän entstammen. Einige Forscher (sogenannte Lumper) stellen noch Fossilien aus dem mittleren Jungpleistozän zu Homo erectus, so dass dieser „Frühmensch“[16] – aus dieser Sicht – fast zwei Millionen Jahre Afrika, Asien und Europa besiedelt hat.
Im April 2020 wurde in der Fachzeitschrift Science das Fossil DNH 134 beschrieben, ein Kinderschädel aus Südafrika, der mit 2,04 bis 1,95 Millionen Jahre älteste bislang bekannte Fund von Homo erectus.[17] Mindestens 1,855 Millionen Jahre alt ist ferner das Hinterhauptbein KNM-ER 2598 aus Kenia.[18] Als jüngster Beleg für Homo erectus gelten die Funde der zunächst als Homo soloensis bezeichneten Fossilien, denen im Jahr 2019 ein Alter von 117.000 bis 108.000 Jahren zugeschrieben wurde.[19]
Diesen Datierungen zufolge lebte Homo erectus zur gleichen Zeit wie Paranthropus boisei. In einer 2024 von Grabungsleiterin Louise Leakey publizierten Studie wurde dies belegt durch rund 1,5 Millionen Jahre alte, zur gleichen Zeit in Koobi Fora (Kenia) entstandene Fußspuren beider Arten.[20]
Von wann bis wann eine fossile Art existierte, kann jedoch in aller Regel nur näherungsweise bestimmt werden. Zum einen ist der Fossilbericht lückenhaft: Es gibt meist nur sehr wenige Belegexemplare für eine fossile Art. Zum anderen weisen die Datierungsmethoden zwar ein bestimmtes Alter aus, dies jedoch mit einer erheblichen Ungenauigkeit; diese Ungenauigkeit bildet dann die äußeren Grenzen bei den „Von-bis-Angaben“ für Lebenszeiten. Alle publizierten Altersangaben sind daher vorläufige Datierungen, die zudem nach dem Fund weiterer Belegexemplare möglicherweise revidiert werden müssen.
Vorkommen
Homo erectus war, wie andere hominine Arten, auf Zugang zu Frischwasser angewiesen; er besiedelte daher vor allem die Umgebung von Flusslandschaften (Galeriewälder) und suchte über Jahrtausende hinweg immer wieder dieselben Tümpel auf. Schon vor rund einer Million Jahre war Homo erectus jedoch auch in der Lage, sich zeitweise in trockenem, halbwüstenartigem Buschland aufzuhalten, was auf ein hohes Maß an ökologischer Flexibilität schließen lässt.[21] Die Autoren einer 2025 publizierte Studie vermuteten daher, dass diese Anpassungsfähigkeit zur Ausdehnung seines geografischen Verbreitungsgebiets beigetragen hat: Homo erectus gilt als die erste Art der Gattung Homo, die sich weit über Afrika hinaus verbreitete.
Heute werden ihm zahlreiche Fossilienfunde zugeordnet, die von ihren Entdeckern eigene Gattungs- und Artnamen erhalten hatten, so zum Beispiel der von Eugène Dubois benannte Anthropopithecus („Java-Mensch“), der von Davidson Black benannte Sinanthropus pekinensis („Peking-Mensch“), der von John T. Robinson benannte Telanthropus capensis („Zielmensch“, ein Fund aus Swartkrans, Südafrika) und der von Camille Arambourg benannte Atlanthropus mauritanicus („Atlas-Mensch“, drei Unterkiefer-Fragmente aus Ternifine bei Muaskar, Algerien; gelegentlich auch: Homo mauritanicus oder Homo erectus mauritanicus).
Zugleich herrschen unter den Paläoanthropologen unterschiedliche Auffassungen über die Abgrenzung des Homo erectus von anderen Arten der Gattung Homo. Seit etwa 1990 werden die ältesten, rund 1,8 bis 1,5 Millionen Jahre alten afrikanischen Funde von Homo erectus – vor allem von einigen US-amerikanischen Paläoanthropologen – als eigene Chronospezies ausgewiesen und als Homo ergaster bezeichnet. Unterschiedliche Auffassungen herrschen auch in Bezug auf jüngere Funde aus dem Mittelpleistozän, die einige Forscher zu Homo erectus stellen, andere aber als Homo rhodesiensis (Funde aus Afrika), Homo antecessor (Funde aus Spanien) oder Homo heidelbergensis (Funde aus Europa) bezeichnen. Möglicherweise gehört auch Homo floresiensis zum Formenkreis des späten Homo erectus.
Die „früheste sicher datierte Anwesenheit von Homininen in Europa“,[22] belegt durch 1,42 Millionen Jahre alte Steinwerkzeuge aus der Fundstätte Korolevo in der Ukraine, wird ebenso Homo erectus zugeschrieben wie die ab 1991 in Georgien entdeckten, 1,85 Millionen Jahre alten Fossilien von Dmanisi.[23]
Anhand genetischer Marker (Alu-Sequenzen) wurde geschätzt, dass die Population von Homo erectus vor 1,2 Millionen Jahren weltweit nur rund 55.000 Individuen umfasste.[24]
Funde aus Asien
Dubois Zuweisung der von ihm in Trinil entdeckten Fossilien zu den Hominini war zunächst äußerst umstritten. Erst nachdem in den 1920er-Jahren die Peking-Menschen und in den 1930er-Jahren von Gustav Heinrich Ralph von Koenigswald auf Java das Fossil Sangiran II sowie weitere, ähnlich alte Funde entdeckt worden waren, galt die Zuordnung der Java-Menschen zu den Ahnen des modernen Menschen als gesichert. Allerdings wurden diese Funde mit jeweils eigenem Artnamen (meist nach ihrem Fundort) benannt, auch wurden zusätzliche Gattungsnamen vorgeschlagen, so dass die Fossilien aus Java u. a. als Homo soloensis, Homo modjokertensis, Pithecanthropus robustus, Pithecanthropus dubius und Meganthropus palaeojavanicus bezeichnet wurden. Besondere Bedeutung erlangten die am Solo-Fluss gefundenen, 14 Individuen zugeordneten Fossilien aus der Nähe des Dorfes Ngandong (Homo soloensis),[25] da es sich bei ihnen vermutlich um die jüngsten Nachweise der Existenz von Homo erectus handelt.[19]
Funde aus Afrika

(Kopie), Senckenbergmuseum
Da sich die Suche nach Vorfahren des modernen Menschen zunächst auf Asien konzentrierte und die ersten afrikanischen Funde der Gattung Australopithecus Jahrzehnte lang nicht als den Hominini zugehörig akzeptiert wurden, gab es in Afrika bis in die 1940er-Jahre kaum eine gezielte Suche nach Fossilien. Die ersten, später dem Homo erectus zugeschriebenen Fossilien – beispielsweise aus Südafrika (Saldanha 1, 1953) und Algerien (Unterkiefer von Ternifine, 1954) – waren Zufallsfunde; sie waren meist schlecht erhalten und erhielten zunächst eigene Gattungsbezeichnungen (zum Beispiel Atlanthropus mauritanicus). Erstmals 1961 wurden auch afrikanische Funde, die seit 1949 bekannten und zuvor als Telanthropus capensis bezeichneten Fossilien aus Swartkrans, als Homo erectus interpretiert.[26] Ab den frühen 1960er-Jahren kamen dann zahlreiche weitere Funde, vor allem aus Kenia, Tansania und Äthiopien, aber auch aus Marokko (Steinbruch Thomas bei Casablanca, 1969) hinzu, darunter das 1960 geborgene fossile Schädeldach OH 9 aus Olduvai und das 1975 am Turkana-See (Koobi Fora) entdeckte Schädeldach KNM-ER 3733, die beide den Funden aus Sangiran („Java-Menschen“) und Zhoukoudian („Peking-Menschen“) ähneln.
Merkmale
Der Körperbau von Homo erectus konnte anhand der zahlreichen Funde in Asien genauer rekonstruiert werden als jener der früheren Hominini-Arten. Es wurden Arm- und Beinknochen von relativ großwüchsigen und von relativ kleinwüchsigen Individuen entdeckt; jedoch konnte bisher nicht geklärt werden, ob dies Ausdruck eines ausgeprägten Geschlechtsdimorphismus oder einer generell großen statistischen Spannweite der Körpergröße ist.[27] Funden aus Afrika wird ferner eine erhebliche Variationsbreite der Knochendicke von Oberschenkelknochen und Becken zugeschrieben.[28]
Körperbau
Charakteristisch für Homo erectus ist demnach der im Vergleich zum modernen Menschen „tonnenförmigere“ und volumenreichere Rumpf[29] sowie sein kräftiges und im Vergleich zu älteren Arten der Hominini größeres Skelett mit besonders dickwandigen Schädelknochen (Schädeldach zwischen 6 und 11 mm) und kräftigen Überaugenwülsten, „über deren Funktion man bis heute rätselt.“[30] Der Schädel ist – von hinten betrachtet – in Relation zu seiner Höhe auffällig breit.
Der Unterkiefer von Homo erectus war im Vergleich zu Homo sapiens breiter und leicht V-förmig. Ein vorspringendes Kinn fehlte. Die Körpergröße wurde in der älteren Fachliteratur auf maximal 1,60 Meter geschätzt, da zunächst aussagekräftiges Skelettmaterial aus dem Bereich des Rumpfes und der Beine fehlte. Funde aus Afrika (Turkana-See) haben später dazu geführt, dass ausgewachsenen Individuen von Homo erectus inzwischen eine Körpergröße von 1,45 bis 1,80 Meter zugeschrieben wird.[31] Das Körpergewicht wird auf 50 bis 60 kg geschätzt.[32] Besonders vollständig erhalten und daher für den gesamten Körperbau besonders aussagekräftig ist das Fossil KNM WT 15000, genannt Nariokotome-Junge.
Das Gehirnvolumen von Homo erectus vergrößerte sich im Verlauf der zwei Millionen Jahre seiner Existenz deutlich; u. a. deshalb variieren die Angaben zu seinem Gehirnvolumen selbst in der gleichen Region sehr stark: von 650 bis 1250 cm³ für ausgewachsene Individuen (zum Vergleich: Homo sapiens 1100 bis 1800 cm³).[33] In der Regel war das Volumen jedoch deutlich größer als das von Homo habilis (ca. 600 bis 700 cm³) oder Homo rudolfensis (ca. 750 cm³). Das Gehirnvolumen der Homo-erectus-Funde aus Sangiran und Trinil wird mit rund 930 cm³ angegeben, das der Peking-Menschen mit rund 1060 cm³, das der Funde aus Ngandong mit rund 1150 cm³.[34]
Einen wichtigen Hinweis auf die Dauer der Kindheit im Vergleich mit dem modernen Menschen lieferte ein 2008 wissenschaftlich beschriebenes, nahezu vollständig erhaltenes Becken eines weiblichen Homo erectus aus Äthiopien, dessen Alter mit 1,4 bis 0,9 Millionen Jahren angegeben wurde.[35] Anders als aufgrund früherer, allerdings weniger aussagekräftiger Beckenfunde von Homo erectus erwartet, ähnelt das fossile Becken (Sammlungsnummer BSN49/P27) in erheblichem Ausmaß noch den von Australopithecus bekannten Gegebenheiten: Der Beckenring ist relativ groß, so dass bei einer Geburt ein schon relativ großer Schädel mit einem relativ weit entwickelten Gehirn hindurch gepasst hat; von der Beckenbreite lässt sich zudem auf einen insgesamt recht breiten Körper schließen. Daraus wurde abgeleitet, dass die Phase der Kindheit, während der das Gehirn zu einer vollen Größe heranwuchs, noch wesentlich kürzer als bei Homo sapiens war, denn der evolutionäre Trend hin zu einer Verlängerung der Kindheitsphase ging einher mit einem verringerten Entwicklungszustand des Gehirns bei der Geburt. Homo erectus wurde dieser Studie zufolge mit rund 34 bis 36 Prozent des Hirnvolumens eines Erwachsenen geboren, was einem Wert zwischen Schimpansen (40 Prozent) und Mensch (rund 28 Prozent) entspricht. Das älteste überlieferte „moderne“ Becken ist rund 100.000 Jahre alt und stammt aus der Skhul-Höhle im heutigen Israel.
Dem einzigen erhaltenen Kinderschädel von Homo erectus („Mojokerto child“; Sammlungsnummer Perning I), der 1936 auf Java entdeckt[36] und 1994 per 39Ar-40Ar-Methode auf 1,8 Millionen Jahre datiert worden war, wurde 2004 ein Lebensalter bei Todeseintritt von ungefähr einem Jahr zugeschrieben. Das Kind hatte zu diesem Zeitpunkt ein Gehirnvolumen von ungefähr 70 bis 90 Prozent eines Erwachsenen; zum Vergleich: Kinder von Homo sapiens haben im Alter von einem Jahr erst ungefähr 50 Prozent des Gehirnvolumens eines Erwachsenen.[37]
Homo erectus verfügte bereits, wie rund 100 bei Ileret (östlich des Turkana-Sees in Kenia) freigelegte, 1,51 bis 1,53 Millionen Jahre alte fossile Fußspuren belegen, sowohl über einen im Wesentlichen dem modernen Menschen entsprechenden Bau der Füße als auch über eine vergleichbare Form der aufrechten, zweibeinigen Fortbewegungsweise.[38] Dies ermöglichte ihm, „größere Entfernungen zurückzulegen und in unterschiedlichere Habitate zu gelangen,“ was wiederum die Voraussetzung dafür gewesen sei, andere Kontinente zu besiedeln.[39] Ausgehend von einer Gruppe von Fußabdrücken schlossen die Forscher zudem zurück auf das Gewicht und hierdurch auf das Geschlecht der Personen. Diese Analysen ergaben „dass mehrere erwachsene Männer der Gruppe angehörten, was auf eine gewisse Toleranz hindeutet, möglicherweise sogar auf eine Kooperation.“[40]
Auch das Herausbilden der dem Homo sapiens anatomisch ähnlichen Schulter und damit der biomechanischen Voraussetzungen für eine Wurftechnik, die die Jagd mit Speeren ermöglichte, wird in die Zeit des frühen Homo erectus vor etwa zwei Millionen Jahren datiert.[41][42] Dies könnte eine wichtige Rolle in der Hominisation gespielt haben.[43]
2007 wurde in der Türkei ein Schädeldach von Homo erectus aus 500.000 Jahre altem Travertin geborgen, das deutliche Merkmale einer durch Tuberkulose ausgelösten Hirnhautentzündung (Leptomeningitis tuberculosa) aufwies.[44]
Ernährung
Unmittelbare fossile Belege für die Ernährungsweise der frühen Vertreter des Homo erectus sind nicht bekannt. Jedoch wird aus den im Vergleich mit den Australopithecinen kleineren Backenzähnen geschlossen, dass ihre Nahrung zumindest teilweise auch Früchte und andere relativ weiche Bestandteile enthielt, häufiger aber Wurzeln, Fleisch und andere kräftiges Kauen erfordernde Stoffe umfasste.[45] Welchen Anteil Jagdbeute an der Ernährung der frühen Homo-erectus-Populationen hatte und welcher Anteil am Fleischkonsum als Aasfresser beschafft wurde, ist jedoch umstritten.[46][47]
Die späten Vertreter von Homo erectus – sie werden jedoch häufig zu Homo heidelbergensis gestellt – können bereits als Jäger und Sammler bezeichnet werden. So werden in der Sammlung von Eugène Dubois im Naturkundemuseum Naturalis in Leiden zahlreiche Muschelschalen verwahrt, die Homo erectus zugeschrieben und in die Zeit vor 540.000 bis 430.000 Jahren datiert wurden.[48] Zahlreiche Schalen wurden mit Hilfe eines spitzen Gegenstands durchbohrt, und zwar an Stellen, die vermutlich dem Öffnen der Schalen von noch lebenden Muscheln dienten.[49] Einer 2020 publizierten Studie zufolge war Homo erectus unter den klimatischen Bedingungen in der heutigen Kalahari vermutlich zu Ausdauerjagden befähigt, die bis zu 5½ Stunden gedauert haben könnten, ohne dass diese Jäger Wasser mit sich führen mussten.[50]
Modellrechnungen kamen zu dem Ergebnis, dass der im Vergleich mit älteren Arten der Hominini schwache Kauapparat mit zurückgebildeter Kaumuskulatur[51] ursächlich mit der Nutzung von Steinwerkzeugen beim Zerkleinern von Nahrung zu erklären ist.[52]
Kultur
_Fundort_Nariokotome,_Kenia,_Rekonstruktion_im_Neanderthal_Museum.jpg)
Homo erectus lebte in jener Epoche, die als Altsteinzeit bezeichnet wird. Schon für seine frühesten Vertreter ist der Gebrauch von Steingerät nachgewiesen. Diese ersten Steinwerkzeuge (Chopper und Chopping Tools) gehören der Kultur des Oldowan an. Seit rund 1,95 Millionen Jahren ist in Afrika die über Faustkeile definierte Kultur des Acheuléen belegt,[53][54] die mit Homo erectus in Verbindung gebracht wird[55] und durch Funde vom Westufer des Turkana-Sees und aus dem südlichen Äthiopien belegt ist. Die Technik der Werkzeugherstellung ist aufwändig zu erlernen; daher ist anzunehmen, dass Homo erectus schon in dieser Epoche über eine ausgeprägte Tradierung des hierfür nötigen Wissens und über die dafür nötigen geistigen Fähigkeiten verfügte. Aus der Fundstelle Dana Aoule North (DAN5) des Fundplatzes Gona (Äthiopien) wurden 1,6 bis 1,5 Millionen Jahre alte Schädelfragmente und – in unmittelbarem Zusammenhang mit diesen – sowohl Steingerät vom Oldowan- als auch vom Acheuléen-Typus geborgen, was darauf hinweist, dass Homo erectus in dieser Epoche und in dieser Region Werkzeuge beider Varianten hergestellt hat.[56]
Ein besonderer Faustkeil-Fund stammt aus Konso (Äthiopien): Dort wurde das bearbeitete Fragment eines 1,4 Millionen Jahre alten Flusspferd-Knochens entdeckt.[57]
Die ältesten in Asien entdeckten und sicher datierten Steinwerkzeuge stammen aus der seit 1863 bekannten Fundstelle Attirampakkam im Nordwesten von Chennai (Südostindien) und gehören zur Kultur des Acheuléen. Für sie wurde mit Hilfe der Aluminium-Beryllium-Methode ein Mindestalter von 1,07 Millionen Jahren errechnet;[58] möglicherweise sind die in unterschiedlich tiefen Fundschichten nachgewiesenen Faustkeile, Cleaver und Schaber sogar bis zu 1,5 Millionen Jahre alt.[59] Zwei einseitig bearbeitete Faustkeile, deren Mindestalter 920.000 Jahre beträgt, wurden bei den Ausgrabungen im Cagayan Valley auf der philippinischen Insel Luzon gefunden.[60] Die ältesten in China entdeckten Faustkeile stammen aus dem Bose-Becken und sind 800.000 Jahre alt.[61] Bemerkenswert ist, dass von den frühen Homo-erectus-Fundstätten auf Java nur sehr primitive Steinwerkzeuge bekannt sind; keines dieser Werkzeuge wurde in dieser Region auf beiden Seiten bearbeitet.[62]
Die ältesten in Europa entdeckten Steinwerkzeuge stammen aus der Sierra de Atapuerca in Spanien und sind 1,2 bis 1,1 Millionen Jahre alt.[63]
Das rund 1,7 Millionen Jahre alte, weibliche Skelett KNM-ER 1808 aus Kenia wies Knochenfehlbildungen auf, die charakteristisch sind für Hypervitaminose A, verursacht durch den Verzehr extrem großer Mengen von Leber. Die Verformungen der Knochen waren so gravierend, dass die Paläoanthropologen davon ausgehen, das Individuum sei die letzten Wochen bis Monate vor seinem Tod vor Schmerzen nahezu bewegungsunfähig gewesen; hieraus wurde gefolgert, dass es vermutlich mit Wasser und Nahrung versorgt wurde – eine frühe Form sozialer Bindung.[64]
Der Beginn der aktiven Nutzung des Feuers durch Homo erectus ist umstritten. Die ältesten gesicherten Feuerstellen, die zweifelsfrei durch Menschen angelegt wurden, stammen aus der Wonderwerk-Höhle in Südafrika und sind rund eine Million Jahre alt.[65][66] Vor rund 790.000 Jahren wurde im heutigen Israel in der Fundstelle Gesher Benot Ya’aqov Feuer genutzt.[67] In Europa stammen die frühesten als gesichert geltenden Feuerstellen aus der Zeit um 400.000 vor heute; entdeckt wurden sie in Schöningen und Beeches Pit (Suffolk, England).[68]
Möglicherweise war Homo erectus vor der Nutzung des Feuers bereits das „Kochen“ möglich. Dies jedenfalls geht aus einer Studie hervor, die im September 2020 publiziert wurde. Ihr zufolge wurden in Schichten der Olduvai-Schlucht im Norden Tansanias, aus denen man 1,7 Millionen Jahre alte Steinwerkzeuge geborgen hatte, Hinweise auf eine Häufung von bestimmten fossilen Molekülen – darunter Lipide – gefunden. Diese Moleküle wurden interpretiert als Biomarker für thermophile, Sulfate-reduzierende Bakterien, wie sie zum Beispiel typischerweise in heißen Quellen, vergleichbar jenen im Yellowstone-Nationalpark, vorkommen. Wörtlich heißt es in der Studie: Solche bis zu 90 Grad heiße Örtlichkeiten „könnten es den frühen Homininen ermöglicht haben, essbare Pflanzen und Fleisch durch Erhitzen zuzubereiten, so dass es in der Evolution des Menschen möglicherweise eine kulturelle Epoche vor dem Feuer gab“ („a prefire stage of human evolution“).[69]
Archäologische Funde, die auf das Herstellen oder Tragen von Kleidung hindeuten könnten, sind nicht bekannt.
Zur Abgrenzung von anderen Arten der Gattung Homo
.png)
.png)
.png)
Homo erectus galt unter Paläoanthropologen Jahrzehnte lang als jene Hominini-Art, die als erste alle drei Kontinente der Alten Welt besiedelte. Seit einigen Jahren wird dieser Konsens jedoch vor allem von US-amerikanischen Forschern infrage gestellt. „Die Kernfrage ist, ob die verschiedenen Homininen-Formen Asiens, Afrikas und Europas wirklich derselben oder aber unterschiedlichen Spezies zuzurechnen sind.“[70] Die aus Afrika, Asien und Europa bekannten Fossilien des Homo erectus weisen nämlich „eine große morphologische Streuung auf, die einige Forscher in mehrere Abstammungslinien aufspalten, während andere sie als eine einzige polytypische Art einstufen,“[71] das heißt – ähnlich der Formenvielfalt bei Homo sapiens – als regionale Varianten einer Art.
Gestützt wird die unter anderem von Berhane Asfaw, Meave Leakey und Tim White vertretene Auffassung, Homo erectus habe zahlreiche regionale Varianten aufgewiesen, durch Schädel- und Oberschenkelfunde auf der Halbinsel Bouri am Awash-Fluss in Äthiopien.[72] Hierzu gehören das Schädeldach BOU-VP-2/66, das am 27. Dezember 1997 geborgen und auf ein Alter von rund einer Million Jahre datiert wurde. Aus diesem Fund wurde ein Schädelvolumen von 995 cm³ abgeleitet sowie – gestützt auch durch die Analyse mehrerer Oberschenkelknochen-Fragmente – eine enge morphologische Nähe zu gleich alten asiatischen Funden.
In einem weiteren Vergleich der Schädelmerkmale asiatischer und afrikanischer Funde wurde in einer 2008 publizierten Studie zwar argumentiert, dass einer Abspaltung der ältesten afrikanischen Funde von Homo erectus als Homo ergaster keine eindeutigen Merkmale zugrunde liegen, dass es jedoch signifikante Unterschiede zwischen einerseits den Funden aus Afrika und den Funden aus Georgien, andererseits aus Asien gebe.[73] Schon 2004 hatte Jeffrey H. Schwartz argumentiert, die morphologischen Unterschiede zwischen einerseits den Funden aus Trinil und Sangiran, andererseits aus Afrika seien so groß, dass deren Zusammenführung unter der Bezeichnung Homo erectus „möglicherweise eher eine historische Episode als eine biologische Realität“ sei.[74]
Homo erectus und Homo ergaster
Ursache für diese sehr unterschiedlichen wissenschaftlichen Sichtweisen ist u. a., dass die Gestalt einiger zu Homo erectus gestellten afrikanischen Fossilien deutlich vom Typusexemplar dieser Art (Trinil II) abweicht.[74] Daraus leitete Bernard Wood 1992 die Berechtigung ab, die bis dahin zu Homo erectus gestellten homininen Fossilien aus der Zeit von vor ca. 1,9 bis 1,4 Mio. Jahren als Homo ergaster abzusondern.[75] Dies hat zur Folge, dass seitdem die afrikanischen Fossilien dieser Epoche einschließlich des Typusexemplars von Homo ergaster, des 1971 von Richard Leakey entdeckten Unterkiefers KNM-ER 992,[76] von einigen Forschern als Homo ergaster, von den meisten Forschern aber weiterhin – wie seit den 1950er-Jahren üblich – als Homo erectus bezeichnet werden.
Tatsächlich veränderte sich der Körperbau von Homo erectus in der Zeit vor 1,8 bis 1,5 Millionen Jahren erheblich, so dass er „sehr abweichend von allem, was vor ihm war“ aussah.[77] Zu den Veränderungen gehörten unter anderem ein Trend hin zu kürzeren Armen, längeren Beinen und einem durch die Ileret-Fußspuren belegten, energiesparenden Abfedern des Auftretens durch Fußgewölbe und großen Zeh. Dagegen ähnelte die Gangart der früheren Hominini, bedingt durch ein nicht oder nur schwach ausgebildetes Fußgewölbe, noch derjenigen der heutigen afrikanischen Menschenaffen.
Der Abtrennung der Homo ergaster zugeschriebenen Fossilien von Homo erectus widersprachen im Jahr 2013 jedoch die Erforscher der Fossilien von Dmanisi (Georgien). Die morphologische Vielfalt dieser sehr gut erhaltenen Funde weise darauf hin, dass die rund 1,8 Millionen Jahre alten, afrikanischen Homo-Fossilien in erster Linie die Variation zwischen lokalen Populationen einer einzigen evolvierenden Abstammungsgruppe widerspiegeln, „die sachgerecht Homo erectus benannt ist.“[23]
Homo erectus und Homo habilis / Homo rudolfensis
Seit seiner Entdeckung Anfang der 1960er-Jahre galt Homo habilis als wahrscheinlichster Vorfahre des Homo erectus. 2007 wurde jedoch ein Oberkiefer-Fragment beschrieben (KNM-ER 42703 von der Fundstelle Ileret, östlich des Turkana-Sees), das Homo habilis zugeordnet und auf ein Alter von 1,44 Millionen Jahren datiert wurde; ferner ein gut erhaltenes Schädeldach, das 1,55 Millionen Jahre alt ist und Homo erectus zugeschrieben wurde. Diesen Datierungen und anderen, älter datierten Funden zufolge existierten Homo habilis und Homo erectus bis zu einer halben Million Jahre nebeneinander. Die Wissenschaftler vermuten, dass beide Arten unterschiedliche ökologische Nischen besetzten und so keine direkten Konkurrenten waren.[78]
Diese Befunde bedeuten zugleich, dass Homo habilis kein Vorfahre des Homo erectus gewesen sein kann; als wahrscheinlichster Vorfahre von Homo ergaster / Homo erectus gilt daher derzeit Homo rudolfensis. Die verwandtschaftlichen Beziehungen von Homo habilis, Homo rudolfensis und Homo ergaster / Homo erectus sind in Fachkreisen allerdings umstritten: Die Körpergröße von Homo erectus, sein verringerter Sexualdimorphismus, die Länge der Gliedmaßen und ganz generell seine Körperproportionen weichen so stark von Homo rudolfensis ab, dass beide Arten primär aufgrund der zeitlichen Aufeinanderfolge ihres Auftretens und weniger aufgrund morphologischer Merkmale als Vor- bzw. Nachfahren erwogen werden.[79]
Homo erectus und Homo heidelbergensis
Die Anfang der 1950er-Jahre noch bescheidene Anzahl von Hominini-Funden aus der Epoche vor 2 Millionen bis 500.000 Jahren hatte dazu geführt, dass all diese Funde der Art Homo erectus zugeordnet wurden. Dies galt auch für den Unterkiefer von Mauer, den sein Erstbeschreiber nach der nahen Universitätsstadt Homo heidelbergensis benannt hatte; diese Bezeichnung blieb jedoch bis in die 1980er-Jahre hinein – wenn überhaupt – allein auf den Unterkiefer von Mauer bezogen. Begründet wurde die Ablehnung einer weiteren Chronospezies damit, dass die stammesgeschichtliche Entwicklung in Europa gleitend von Homo erectus zum Neandertaler verlief; jede weitere Grenzziehung sei willkürlich, weswegen die Paläoanthropologen auf die Verwendung des Artnamens Homo heidelbergensis verzichteten. Selbst der Unterkiefer von Mauer wurde daher als lokale (europäische) Spätform des Homo erectus ausgewiesen, als Homo erectus heidelbergensis. Ähnlich argumentieren die französischen Entdecker des Homo erectus tautavelensis, die osteuropäischen Erforscher des Homo erectus bilzingslebensis und Alfred Czarnetzki, der Bearbeiter des Homo erectus reilingensis aus Reilingen.
Eine zweite Gruppe von Forschern bezeichnet heute zwar die bis zu 1,8 Millionen Jahre alten afrikanischen Fossilien als Homo ergaster oder Homo erectus und leitet aus diesen die Besiedelung Europas und Asiens ab, benennt die europäischen Nachfahren dieser afrikanischen Auswanderer aber als Homo heidelbergensis. Ihren Publikationen zufolge verläuft die Entwicklung zunächst in Afrika von Homo ergaster zu Homo erectus und danach in Europa über Homo heidelbergensis zum Neandertaler. Bernard Wood begründete die Zuordnung der europäischen Fossilien zu einer eigenen Chronospezies bereits 1984 damit, dass die Ähnlichkeit der asiatischen und der afrikanische Funde zwar „eindeutig und widerspruchsfrei“ („clear and unambiguous“) sei, die Übereinstimmung der europäischen Funde hingegen nicht überzeugend dargelegt wurde.[80]
Eine dritte Gruppierung von Forschern fordert schließlich eine noch weitergehende Änderung der Zuweisung von afrikanischen Fossilien zu einer Art der Gattung Homo. Von ihnen „wurde dieses umfassende Taxon aus chronologischen und geographischen Erwägungen aufgespalten“, Homo erectus wird von diesen Forschern seitdem „als Vertreter einer spezifisch ostasiatischen Stammlinie“ ausgewiesen.[81] Die ältesten, von anderen Wissenschaftlern weiterhin zu Homo erectus gestellten afrikanischen Fossilien werden von diesen Forschern als Homo ergaster bezeichnet, die jüngeren, unmittelbar an Homo ergaster anschließenden Fossilien aber als Homo heidelbergensis. Dieser Konvention zufolge entwickelte sich Homo ergaster bereits in Afrika zu Homo heidelbergensis fort, während aus Afrika nach Asien ausgewanderte Gruppen von Homo ergaster sich in Asien zu Homo erectus entwickelten.
Da sich die Paläoanthropologen bisher auf keine gemeinsame Konvention verständigen konnten, werden seit einigen Jahren – je nach Vorliebe der einzelnen Autoren – bestimmte Fossilien in Fachveröffentlichungen mal zur einen, mal zur anderen Art gestellt. Es wurde sogar eingewandt, dass selbst die Begrenzung des Homo erectus auf Asien noch viel zu unterschiedlich aussehende afrikanische Fossilien zu einer Art bündele. Die britische Paläoanthropologin Leslie Aiello wurde beispielsweise in der Fachzeitschrift Science zitiert, beim afrikanischen Homo heidelbergensis handele es sich um ein „Mülleimer-Taxon“; sie schlug vor, die Art Homo heidelbergensis europäischen Fossilien vorzubehalten und die afrikanischen Nachkommen von Homo ergaster zu einer bislang noch nicht benannten neuen Art zu erheben.[82]
Homo erectus, Homo rhodesiensis und Homo antecessor
Der Vorschlag jener Gruppierung von Forschern, die unmittelbaren afrikanischen Nachfahren von Homo ergaster als Homo heidelbergensis auszuweisen, hat zur Folge, dass der so definierte afrikanische Homo heidelbergensis als Vorfahre des Homo sapiens zu gelten hat.[83] Wird hingegen in einer Fachveröffentlichung die alternative Lesart zugrunde gelegt, der zufolge Homo heidelbergensis eine erst in Europa aus Homo erectus entstandene Vorläuferart der Neandertaler ist, dann gibt es zwischen Homo heidelbergensis und Homo sapiens keine derart enge verwandtschaftliche Beziehung.
Vollends verwirrend wurde die Zergliederung des afrikanischen Homo erectus, als vorgeschlagen wurde, die in Afrika entdeckten unmittelbaren Vorfahren des Homo sapiens vom afrikanischen Homo heidelbergensis abzuspalten und als Homo rhodesiensis zu bezeichnen – ein Vorschlag, der sich bisher international nicht durchgesetzt hat. Diese rund 200.000 Jahre alten afrikanischen Fossilien werden heute gleichwohl – abhängig von der jeweiligen Forschergruppe – als früher Homo sapiens, archaischer Homo sapiens, Homo rhodesiensis, später Homo heidelbergensis oder später Homo erectus bezeichnet.
Spanische Forscher haben die Vielfalt der Benennungen 1994 schließlich um eine weitere bereichert: um den bisher nur aus Spanien bekannten Homo antecessor, dessen Einstufung als eigenständige Art allerdings umstritten ist. Einige spanische Forscher vertreten in ihren Publikationen zwar die Auffassung, Homo antecessor habe sich in Afrika aus Homo erectus entwickelt, sei dann in Europa eingewandert und habe sich hier zu Homo heidelbergensis fortentwickelt. Andere Forscher ordnen den älteren Teil der spanischen Fossilien hingegen Homo erectus zu, der sich in Europa zu Homo heidelbergensis fortentwickelt habe.
Historischer Hintergrund
Die Vielfalt unterschiedlicher Abgrenzungen hat ihren Ursprung unter anderem in der Argumentation Ernst Mayrs, die Vorfahren des Homo sapiens hätten einen ähnlich variablen Körperbau besessen wie die Jetzt-Menschen[8] und es sei unangebracht, die Unterschiede zwischen den Fossilien von unterschiedlichen Fundorten zu betonen. Er entwickelte jedoch keinen Katalog der artspezifischen Merkmale. Stattdessen bezeichnete er die ältesten, aus Südafrika stammenden Funde mit einer geographischen Zuordnung als Homo transvaalensis (nach den Fundorten in der Provinz Transvaal, heute lautet die Bezeichnung Australopithecus africanus); zwischen diese Art und Homo sapiens platzierte er einzig Homo erectus. Somit wurden „alle Funde des frühen und mittleren Pleistozäns mit einem Hirnvolumen zwischen ungefähr 800 und 1200 Kubikzentimetern einer einzigen Art zugeordnet“.[84] Auch Mayrs Festlegungen, die unter anderem eine geradlinige Transformation der älteren Art zur nächstjüngeren unterstellten, wurden nicht von klaren Beschreibungen der diese drei Arten unterscheidenden Merkmale (Diagnosen) unterfüttert. Deshalb und in dem Maße, in dem sich später die Auffassung durchsetzte, dass es sehr wohl auch ausgestorbene „Seitenäste“ im Stammbaum des Menschen gab, neigten etliche Forscher dazu, die Unterschiede der Fossilien zu betonen und in der Folge die Namensvielfalt erneut zu vergrößern. Zudem wurde wiederholt argumentiert, dass das Typusexemplar von Homo erectus, Trinil II, einige sehr spezielle „abgeleitete“ Merkmale aufweise, die es „ausschließen“, dass das Fossil „der Vorfahre von irgend einer anderen Homo-Art einschließlich Homo sapiens“ ist.[85] Ähnlich unklar ist allerdings auch die Zuordnung der Fossilien zu Homo ergaster; hier besteht der Einwand, dass die Anatomie von dessen Typusexemplar, des Unterkiefer-Fragments KNM-ER 992, erheblich abweiche vom ebenfalls zu Homo ergaster gestellten Unterkiefer des Nariokotome-Jungen.
Ian Tattersall zufolge wurden beispielsweise 1979 das Schädeldach OH 9 („Chellean man“) aus Tansania,[86] 1998 der Schädel UA 31 aus Buia[87] (Afar, Eritrea), 2002 das Schädeldach BOU-VP-2/66 („Daka Calvaria“) vom Fundort Bouri in Äthiopien[88] und 2003 das Schädelfragment aus Olorgesailie (Kenia) Homo erectus zugeordnet, obwohl charakteristische Merkmale dieser vier Funde sowohl vom Typusexemplar der Art aus Java als auch voneinander abweichen.[89] In einer Übersichtsarbeit zur Herkunft des Homo sapiens kamen Jeffrey H. Schwartz und Ian Tattersall daher zu folgendem Ergebnis:
- „Jedes Bemühen, den Ursprung von H. sapiens direkt auf H. erectus zurückzuführen, hängt davon ab, welche Fundstücke man der letztgenannten Art zurechnet, was sich dann direkt auf jene Szenarien auswirkt, wann, wo und, abhängig vom Ausmaß der Spekulation, wie H. erectus den H. sapiens hervorgebracht hat.“[90]
Literatur
- Bernard Wood: Birth of Homo erectus. In: Evolutionary Anthropology. Band 29, Nr. 6, 2020, S. 293–298, doi:10.1002/evan.21873, Volltext.
- Jean-Jacques Hublin: Paleoanthropology: Homo erectus and the Limits of a Paleontological Species. In: Current Biology. Band 24, Nr. 2, 2014, S. R82–R84, doi:10.1016/j.cub.2013.12.006.
- Andrew Kramer: Human taxonomic diversity in the pleistocene: Does Homo erectus represent multiple hominid species? In: American Journal of Physical Anthropology. Band 91, Nr. 2, 1993, S. 161–171, doi:10.1002/ajpa.1330910203.
- Philip Rightmire: The Evolution of Homo erectus. Comparative Anatomical Studies of an Extinct Human Species. Cambridge University Press, 1990, ISBN 978-0-511-52567-4, doi:10.1017/CBO9780511525674.
- Bernard Wood: The origin of Homo erectus. In: Courier Forschungsinstitut Senckenberg. Band 69, 1984, S. 99–111.
- Peter Andrews: An alternative interpretation of the characters used to define Homo erectus. In: Courier Forschungsinstitut Senckenberg. Band 69, 1984, S. 167–175.
- Phillip Tobias und Gustav Heinrich Ralph von Koenigswald: A Comparison Between the Olduvai Hominines and those of Java and some Implications for Hominid Phylogeny. In: Nature. Band 204, 1964, S. 515–518, doi:10.1038/204515a0.
Erstbeschreibung:
- Eugène Dubois: Paleontologische onderzoekingen op Java. Verslag van het Mijnwezen, 3. Qu. 1892, S. 10–14.
- Eugène Dubois: Paleontological Investigations on Java. In: William Eric Meikle, Sue Taylor Parker (Hrsg.): Naming our Ancestors. An Anthology of Hominid Taxonomy. Waveland Press, Prospect Heights (Illinois) 1994, S. 37–40, ISBN 0-88133-799-4. (Übersetzung ins Englische durch den Berkeley Scientific Translation Service).
Siehe auch
Weblinks
- Homo erectus: Der unfassbare Frühmensch. Auf: spektrum.de vom 24. Februar 2021.
- Climate played a crucial role in human migration from Africa. Zusammenfassung einer Studie vom 9. Oktober 2023.
- Übersicht zu den Funden von Homo erectus in Asien, Afrika und Europa (englisch, Stand: 2004) ( vom 9. November 2004 im Internet Archive)
- Übersicht zur Entdeckungsgeschichte und zu anatomischen Merkmalen (englisch, Stand: 2005 ( vom 18. Januar 2006 im Internet Archive)
- Bebilderte Übersicht zu den asiatischen Funden von Homo erectus (englisch, Stand: 2001) ( vom 10. Oktober 2012 im Internet Archive)
Belege
- ↑ Richard Leakey: The origin of humankind. Phoenix, a division of Orion Books Ltd., 1995, S. XIV
- ↑ Charles Darwin: The Descent of Man, and Selection in Relation to Sex. John Murray, London 1871, Band 1, S. 199: „In each great region of the world the living mammals are closely related to the extinct species of the same region. It is therefore probable that Africa was formerly inhabited by extinct apes closely allied to the gorilla and chimpanzee; and as these two species are now man’s nearest allies, it is somewhat more probable that our early progenitors lived on the African continent than elsewhere.“
- ↑ Bernard Wood: Fifty years after Homo habilis. In: Nature. Band 508, Nr. 7494, 2014, S. 31–33, doi:10.1038/508031a
- ↑ Eugène Dubois publizierte seine Forschungsergebnisse anfangs quartalsweise in den Berichten des Minenwesens („Verslag van het Mijnwezen“) der niederländischen Kolonialverwaltung; die Benennung erfolgte im Bericht für das 3. Quartal 1892, (erschienen 1893), S. 10–14; vergl. dazu: Aleš Hrdlička: The skeletal remains of early man. Smithsonian Institution, 1930; Volltext.
- ↑ Eugène Dubois: Pithecanthropus erectus: eine menschenähnliche Übergangsform von Java. Landes-Druckerei, Batavia 1894
- ↑ Stephanie Müller et al.: Sangiran II. Eine Schädelkalotte und ihre wissenschaftliche Deutung. In: Natur und Museum. Band 138, 2008, S. 30
- ↑ Theodosius Dobzhansky: On species and races of living and fossil man. In: American Journal of Physical Anthropology. Band 2, Nr. 3, 1944, S. 251–265, doi:10.1002/ajpa.1330020303
- ↑ a b Ernst Mayr: Taxonomic categories in fossil hominids. Cold Spring Harbor Symposia on Quantitative Biology 15, 1950, S. 109–118; Exzerpt. Nachdruck in: W. Eric Meikle, Sue Taylor Parker: Naming our Ancestors. An Anthology of Hominid Taxonomy. Waveland Press, Prospect Heights (Illinois) 1994, ISBN 0-88133-799-4, S. 152–170.
- ↑ Gary J. Sawyer, Viktor Deak: Der lange Weg zum Menschen. Lebensbilder aus 7 Millionen Jahren Evolution. Spektrum Akademischer Verlag, Heidelberg 2008, S. 117 und 119
- ↑ Gary J. Sawyer und Viktor Deak, Der lange Weg zum Menschen, S. 121.
- ↑ Michael Herbert Day und Theya I. Molleson: The Trinil femora. In: M. H. Day (Hrsg.): Human Evolution. Symposia of the Society for the Study of Human Biology. Band 11, Taylor & Francis, London 1973, S. 127–154.
- ↑ Daniel John Cunningham: Dr. Dubois' So-Called Missing Link. In: Nature. Band 51, 1895, S. 428–429, doi:10.1038/051428a0.
- ↑ Bert Theunissen: Eugène Dubois and the Ape-Man from Java. The History of the First ‘Missing Link’ and Its Discoverer. Springer 1989, ISBN 978-1-55608-081-4, doi:10.1007/978-94-009-2209-9.
- ↑ W. Eric Meikle und Sue Taylor Parker (Hrsg.): Naming our Ancestors. An Anthology of Hominid Taxonomy. Waveland Press, Prospect Heights (Illinois) 1994, S. 36, ISBN 0-88133-799-4
- ↑ Eduard Pop, Sofwan Noerwidi und Fred Spoor: Naming Homo erectus: A review. In: Journal of Human Evolution. Band 190, 2024, 103516, doi:10.1016/j.jhevol.2024.103516.
- ↑ In Grzimeks Tierleben wird Homo erectus als „Frühmensch“ bezeichnet; siehe dazu die „Systematische Übersicht“ ebd., Band 11 (= Säugetiere 2), dtv, 1979, S. 508. Die Gattung Australopithecus wird hier erläuternd als „Gattung Vormensch“ bezeichnet.
- ↑ Andy I. R. Herries et al.: Contemporaneity of Australopithecus, Paranthropus, and early Homo erectus in South Africa. In: Science. Band 368, Nr. 6486, 2020, eaaw7293, doi:10.1126/science.aaw7293.
Our direct human ancestor Homo erectus is older than we thought. Auf: eurekalert.org vom 2. April 2020. - ↑ Ashley S. Hammond et al.: New hominin remains and revised context from the earliest Homo erectus locality in East Turkana, Kenya. In: Nature Communications. Band 12, Artikel-Nr. 1939, 2021, doi:10.1038/s41467-021-22208-x.
Study cements age and location of hotly debated skull from early human Homo erectus. Auf: eurekalert.org vom 13. April 2021. - ↑ a b Yan Rizal et al.: Last appearance of Homo erectus at Ngandong, Java, 117,000–108,000 years ago. In: Nature. Band 577, 2020, S. 381–385, doi:10.1038/s41586-019-1863-2.
Researchers determine age for last known settlement by a direct ancestor to modern humans. Auf: eurekalert.org vom 18. Dezember 2019. - ↑ Kevin G. Hatala et al.: Footprint evidence for locomotor diversity and shared habitats among early Pleistocene hominins. In: Science. Band 386, Nr. 6725, 2024, S. 1004–1010, doi:10.1126/science.ado5275.
Menschenvorfahren schritten Seit' an Seit'. Auf: spektrum.de vom 28. November 2024. - ↑ Julio Mercader et al.: Homo erectus adapted to steppe-desert climate extremes one million years ago. In: Communications Earth & Environment. Band 6, Nr. 1, 2025, doi:10.1038/s43247-024-01919-1.
Human ancestor thrived longer in harsher conditions than previous estimates. Auf: eurekalert.org vom 16. Januar 2025. - ↑ Roman Garba et al.: East-to-west human dispersal into Europe 1.4 million years ago. In: Nature. Online-Vorabveröffentlichung vom 6. März 2024, doi:10.1038/s41586-024-07151-3.
Ältestes Zeugnis von Frühmenschen in Europa. Auf: scinexx.de vom 7. März 2024. - ↑ a b David Lordkipanidze et al.: A Complete Skull from Dmanisi, Georgia, and the Evolutionary Biology of Early Homo. In: Science. Band 342, Nr. 6156, 2013, S. 126–131, doi:10.1126/science.1238484
- ↑ Chad D. Huff et al.: Mobile elements reveal small population size in the ancient ancestors of Homo sapiens. In: PNAS. Band 107, Nr. 5, 2010, S. 2147–2152, doi:10.1073/pnas.0909000107. Zu beachten ist bei dieser Studie, dass die effektive Populationsgröße stets kleiner ist als die Zahl der Individuen.
- ↑ W. F. F. Oppennoorth: Homo (Javanthropus) soloensis: een plistocene mensch van Java. In: Wetenschappelijke medeligen Dienst van den Mijnbrouw in Nederlandsch-Indië. Band 20, 1932, S. 49 ff.
- ↑ Winfried Henke, Hartmut Rothe: Stammesgeschichte des Menschen. Springer Verlag, 1999, S. 199
- ↑ Ann Gibbons: Human Ancestor Caught in the Midst of a Makeover. In: Science. Band 328, Nr. 5977, 2010, S. 413, doi:10.1126/science.328.5977.413
- ↑ Carol V. Ward et al.: Associated ilium and femur from Koobi Fora, Kenya, and postcranial diversity in early Homo. In: Journal of Human Evolution. Band 81, 2015, S. 48–67, doi:10.1016/j.jhevol.2015.01.005
- ↑ Robert G. Franciscus, Steven E. Churchill: The costal skeleton of Shanidar 3 and a reappraisal of Neandertal thoracic morphology. In: Journal of Human Evolution. Band 42, 2002, S. 303–356, doi:10.1006/jhev.2001.0528
- ↑ Friedemann Schrenk: Die Frühzeit des Menschen. Der Weg zum Homo sapiens. C. H. Beck, 5., vollständig neubearbeitete und ergänzte Auflage, München 2008, S. 93 (C.H.Beck Wissen), ISBN 978-3-406-57703-1
- ↑ David Lordkipanidze et al.: Postcranial evidence from early Homo from Dmanisi, Georgia. In: Nature. Band 449, 2007, S. 305–310; doi:10.1038/nature06134
- ↑ Thorolf Hardt, Bernd Herkner und Ulrike Menz: Safari zum Urmenschen. Schweizerbart’sche Verlagsbuchhandlung, Stuttgart 2009, S. 122, ISBN 978-3-510-61395-3
- ↑ Russell L. Ciochon, E. Arthur Bettis III: Asian Homo erectus converges in time. In: Nature. Band 458, 2009, S. 153–154, doi:10.1038/458153a
- ↑ Gary J. Sawyer, Viktor Deak, S. 85 (H. habilis), S. 79 (H. rudolfensis), S. 115 und 124 (H. erectus)
- ↑ Scott W. Simpson et al.: A Female Homo erectus Pelvis from Gona, Ethiopia. In: Science. Band 322, Nr. 5904, 2008, S. 1089–1092, doi:10.1126/science.1163592
- ↑ O. Frank Huffman, Pat Shipman et al.: Historical Evidence of the 1936 Mojokerto Skull Discovery, East Java. In: Journal of Human Evolution. Band 18, Nr. 4, 2005, S. 321–364, doi:10.1016/j.jhevol.2004.09.001.
- ↑ Hélène Coqueugniot et al.: Early brain growth in Homo erectus and implications for cognitive ability. In: Nature. Band 431, 2004, S. 299–302, doi:10.1038/nature02852
- ↑ „The Ileret prints show that by 1.5 Ma, hominins had evolved an essentially modern human foot function and style of bipedal locomotion.“ Matthew R. Bennett et al.: Early Hominin Foot Morphology Based on 1.5-Million-Year-Old Footprints from Ileret, Kenya. In: Science. Band 323, 2009, S. 1197–1201; doi:10.1126/science.1168132
- ↑ Jack Harris, Rutgers University, Co-Autor der Publikation zu den Ileret-Fußspuren, zitiert in: Robert Adler: Fossil footprints reveal modern walk. In: New Scientist vom 7. März 2009, S. 10 und Online-Version vom 26. Februar 2009.
- ↑ Homo erectus ging wie wir. Auf: mpg.de vom 12. Juli 2016.
Kevin G. Hatala et al.: Footprints reveal direct evidence of group behavior and locomotion in Homo erectus. In. Scientific Reports. Band 6, Artikel-Nr. 28766, 2016, doi:10.1038/srep28766. - ↑ Jan Dönges: Wie der Mensch zu seinem einzigartigen Wurftalent kam. spektrum.de vom 27. Juni 2013 (mit Video), abgerufen am 28. März 2022.
- ↑ Martin Vieweg: Ein buchstäblich großer Wurf. Auf: wissenschaft.de vom 26. Juni 2013.
- ↑ Neil T. Roach, Madhusudhan Venkadesan, Michael J. Rainbow & Daniel E. Lieberman: Elastic energy storage in the shoulder and the evolution of high-speed throwing in Homo. In: Nature. Band 498, 2013, S. 483–486, doi:10.1038/nature12267
- ↑ John Kappelman et al.: First Homo erectus from Turkey and implications for migrations into temperate Eurasia. In: American Journal of Physical Anthropology. Band 135, 2008, S. 110–116, doi:10.1002/ajpa.20739.
„Ältester Tuberkulose-Nachweis beim Menschen?“ Auf: idw-online.de vom 23. April 2012. - ↑ Peter S. Ungar et al.: Dental microwear and diets of African early Homo. In: Journal of Human Evolution. Band 50, 2006, S. 78–95, doi:10.1016/j.jhevol.2005.08.007; vergl. dazu: Homo erectus ate crunchy food. Auf: abc.net.au vom 22. November 2005.
- ↑ James F. O’Connell, Kristen Hawkes, Karen Lupo, N. G. Blurton Jones: Male strategies and Plio-Pleistocene archaeology. In: Journal of Human Evolution. Band 43, 2002, S. 831–872, doi:10.1006/jhev.2002.0604, Volltext (PDF; 929 kB) ( vom 8. Juli 2014 im Internet Archive); vergl. dazu „Frauen sorgten für Ernährung“: Überlegungen zur Ernährung von Homo erectus
- ↑ W. Andrew Barr et al.: No sustained increase in zooarchaeological evidence for carnivory after the appearance of Homo erectus. In: PNAS. Band 119, Nr. 5, e2115540119; doi:10.1073/pnas.2115540119.
- ↑ Etchings on a 500,000-year-old shell appear to have been made by human ancestor. Auf: sciencemag.org vom 3. Dezember 2014.
- ↑ Josephine C. A. Joordens et al.: Homo erectus at Trinil on Java used shells for tool production and engraving. In: Nature. Band 518, Nr. 7538, 2015, S. 228–231, doi:10.1038/nature13962
- ↑ Martin Hora et al.: Dehydration and persistence hunting in Homo erectus. In: Journal of Human Evolution. Band 138, 2020, 102682, doi:10.1016/j.jhevol.2019.102682.
- ↑ Hansell H. Stedman et al.: Myosin gene mutation correlates with anatomical changes in the human lineage. In: Nature. Band 428, 2004, S. 415–418, doi:10.1038/nature02358
- ↑ Katherine D. Zink und Daniel E. Lieberman: Impact of meat and Lower Palaeolithic food processing techniques on chewing in humans. In: Nature. Band 531, 2016, S. 500–503, doi:10.1038/nature16990
- ↑ Margherita Mussi et al.: Early Homo erectus lived at high altitudes and produced both Oldowan and Acheulean tools. In: Science. Band 382, Nr. 6671, 2023, S. 713–718, doi:10.1126/science.add9115.
About 2 million years ago, Homo Eerectus lived at high altitudes and produced both Oldowan and Acheulean tools. Auf: eurekalert.org vom 12. Oktober 2023. - ↑ Christopher J. Lepre et al.: An earlier origin for the Acheulian. In: Nature. Band 477, 2011, S. 82–85, doi:10.1038/nature10372
- ↑ Yonas Beyene et al.: The characteristics and chronology of the earliest Acheulean at Konso, Ethiopia. In: PNAS. Band 110, Nr. 5, 2013, S. 1584–1591, doi:10.1073/pnas.1221285110
- ↑ Sileshi Semaw et al.: Co-occurrence of Acheulian and Oldowan artifacts with Homo erectus cranial fossils from Gona, Afar, Ethiopia. In: Science Advances. Band 6, Nr. 10, 2020, eaaw4694, doi:10.1126/sciadv.aaw4694.
Homo erectus used two different kinds of stone tools. Auf: newscientist.com vom 5. März 2020. - ↑ Katsuhiro Sano et al.: A 1.4-million-year-old bone handaxe from Konso, Ethiopia, shows advanced tool technology in the early Acheulean. In: PNAS. Band 117, Nr. 31, 2020, S. 18393–18400, doi:10.1073/pnas.2006370117.
Ran Barkai: Lower Paleolithic bone handaxes and chopsticks: Tools and symbols? In: PNAS. Band 117, Nr. 49, 2020, S. 30892–30893, doi:10.1073/pnas.2016482117.
Gen Suwa, Berhane Asfaw et al.: Reply to Barkai: Implications of the Konso bone handaxe. In: PNAS. Band 117, Nr. 49, 2020, S. 30894–30895, doi:10.1073/pnas.2018084117. - ↑ Shanti Pappu et al.: Early Pleistocene Presence of Acheulian Hominins in South India. In: Science. Band 331, Nr. 6024, 2011, S. 1596–1599, doi:10.1126/science.1200183
- ↑ Robin Dennell: An Earlier Acheulian Arrival in South Asia. In: Science. Band 331, Nr. 6024, 2011, S. 1532–1533, doi:10.1126/science.1203806
- ↑ Paleolithic Archaeological Sites in Cagayan Valley., abgerufen am 3. August 2013: Beschreibung der Funde auf Luzon auf der Website der UNESCO
- ↑ Hou Yamei: Mid-Pleistocene Acheulean-like Stone Technology of the Bose Basin, South China. In: Science. Band 287, 2000, S. 1622–1626, doi:10.1126/science.287.5458.1622, Kurzfassung (auf Englisch) ( vom 13. Februar 2008 im Internet Archive)
- ↑ G. J. Sawyer, Viktor Deak: Der lange Weg zum Menschen, S. 118
- ↑ Eudald Carbonell et al.: The first hominin of Europe. In: Nature. Band 452, 2008, S. 465–469, doi:10.1038/nature06815
- ↑ Alan Walker, Michael R. Zimmerman und Richard Leakey: A possible case of hypervitaminosis A in Homo erectus. In: Nature. Band 296, 1982, S. 248–250, doi:10.1038/296248a0
- ↑ F. Berna, P. Goldberg, L. K. Horwitz, J. Brink, S. Holt, M. Bamford, M. Chazan: Microstratigraphic evidence of in situ fire in the Acheulean strata of Wonderwerk Cave, Northern Cape province, South Africa. In: Proceedings of the National Academy of Sciences. 109. Jahrgang, Nr. 20, 2012, ISSN 0027-8424, S. E1215–E1220, doi:10.1073/pnas.1117620109.
- ↑ Eine Million Jahre alte Brandspuren: Schon Homo erectus spielte mit dem Feuer. Auf: spiegel.de vom 3. April 2012.
- ↑ Naama Goren-Inbar et al.: Evidence of Hominin Control of Fire at Gesher Benot Ya`aqov, Israel. In: Science. Band 304, 2004, S. 725–727, doi:10.1126/science.1095443
- ↑ Wil Roebroeks, Paola Villa: On the earliest evidence for habitual use of fire in Europe. In: PNAS. Band 108, Nr. 13, 2011, S. 5209–5214, doi:10.1073/pnas.1018116108
- ↑ Ainara Sistiaga et al.: Microbial biomarkers reveal a hydrothermally active landscape at Olduvai Gorge at the dawn of the Acheulean, 1.7 Ma. In: PNAS. Online-Vorabveröffentlichung vom 15. September 2020, doi:10.1073/pnas.2004532117.
Did our early ancestors boil their food in hot springs? Auf: mit.edu vom 15. September 2020. - ↑ Winfried Henke, Hartmut Rothe: Stammesgeschichte des Menschen, S. 205
- ↑ Richard Potts et al.: Small Mid-Pleistocene Hominin Associated with East African Acheulean Technology. In: Science. Band 305, 2004, S. 75–78, doi:10.1126/science.1097661
- ↑ Berhane Asfaw et al.: Remains of Homo erectus from Bouri, Middle Awash, Ethiopia. In: Nature. Band 416, 2002, S. 317–320, doi:10.1038/416317a. Vergleiche dazu. Homo erectus: Weltweiter Urahne des Menschen. Auf: orf.at, 2002
- ↑ Karen L. Baab: The taxonomic implications of cranial shape variation in Homo erectus. In: Journal of Human Evolution. Band 54, Nr. 6, 2008, S. 827–847, doi:10.1016/j.jhevol.2007.11.003, Volltext (PDF; 993 kB)
- ↑ a b Jeffrey H. Schwartz: Getting to Know Homo erectus. In: Science, Band 305, 2004, S. 53–54; doi:10.1126/science.1099989
- ↑ Bernard Wood: Origin and evolution of the genus Homo. In: Nature. Band 355, 1992, S. 783–790, doi:10.1038/355783a0; vergl. dazu auch: Bernard Wood: Early hominid species and speciation. In: Journal of Human Evolution, Band 22, Nr. 4–5, 1992, S. 351–365, doi:10.1016/0047-2484(92)90065-H
- ↑ Unterkiefer KNM-ER 992, Abbildung ( vom 6. November 2007 im Internet Archive)
- ↑ Matthew Bennett, Bournemouth University, Hauptautor der Publikation zu den Ileret-Fußspuren, zitiert in: Robert Adler: Fossil footprints reveal modern walk. In: New Scientist vom 7. März 2009, S. 10 und Online-Version vom 26. Februar 2009
- ↑ Fred Spoor et al.: Implications of new early Homo fossils from Ileret, east of Lake Turkana, Kenya. In: Nature. Band 448, 2007, S. 688–691; doi:10.1038/nature05986
vergl. dazu „Fossilien lassen menschlichen Stammbaum wackeln.“ – Meldung auf spiegel.de vom 9. August 2007 - ↑ Robin Dennell, Wil Roebroeks: An Asian perspective on early human dispersal from Africa. In: Nature. Band 438, 2005, S. 1099–1104, doi:10.1038/nature04259
- ↑ Bernard Wood: The origin of Homo erectus. In: Cour. Forsch. Inst. Senckenberg. Band 69, 1984, S. 99–111.
- ↑ Robert Foley: Menschen vor Homo sapiens. Wie und warum unsere Art sich durchsetzte. Jan Thorbecke Verlag, 2000, S. 153.
- ↑ Ann Gibbons: A new face for human ancestors. In: Science. Band 276, 1997, S. 1331–1333, doi:10.1126/science.276.5317.1331.
- ↑ Chris Stringer: The status of Homo heidelbergensis (Schoetensack 1908). In: Evolutionary Anthropology: Issues, News, and Reviews. Band 21, Nr. 3, 2012, S. 101–107, doi:10.1002/evan.21311
- ↑ Peter Andrews: An alternative interpretation of the characters used to define Homo erectus. In: Cour. Forsch. Inst. Senckenberg. Band 69, 1984, S. 167–175
- ↑ Jeffrey H. Schwartz und Ian Tattersall: Fossil evidence for the origin of Homo sapiens. In: American Journal of Physical Anthropology. Band 143, Supplement 51 (= Yearbook of Physical Anthropology), 2010, S. 94–121 (hier S. 99), doi:10.1002/ajpa.21443
- ↑ G. Philip Rightmire: Cranial remains of Homo erectus from Beds II and IV, Olduvai Gorge, Tanzania. In: American Journal of Physical Anthropology. Band 51, Nr. 1, 1979, S. 99–115, doi:10.1002/ajpa.1330510113
- ↑ Ernesto Abbate et al.: A one-million-year-old Homo cranium from the Danakil (Afar) Depression of Eritrea. In: Nature. Band 393, 1998, S. 458–460, doi:10.1038/30954
Der Schädel aus dem Höllenloch der Schöpfung. Auf: welt.de vom 26. Mai 2008 - ↑ Berhane Asfaw, W. Henry Gilbert, Yonas Beyene et al.: Remains of Homo erectus from Bouri, Middle Awash, Ethiopia. In: Nature. Band 416, 2002, S. 317–320, doi:10.1038/416317a
Daka Homo erectus. Abbildung auf leakeyfoundation.org 15. August 2014 - ↑ Ian Tattersall: Masters of the Planet. The Search for Our Human Origins. Palgrave Macmillan, New York 2012, S. 130, ISBN 978-0-230-10875-2
- ↑ Jeffrey H. Schwartz und Ian Tattersall: Fossil evidence for the origin of Homo sapiens, S. 100