
 French
French Deutsch
DeutschMosasaurier – Wikipedia
| Mosasaurier | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
 Hainosaurus in einer Lebendrekonstruktion. Dieser Tylosaurine aus der Oberkreide wurde bis zu 17 Meter lang. | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Oberkreide (Cenomanium bis Maastrichtium) | ||||||||||||
| 100,5 bis 66 Mio. Jahre | ||||||||||||
| Fundorte | ||||||||||||
| ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Mosasauridae | ||||||||||||
| Gervais, 1853 | ||||||||||||
Die Mosasaurier (Mosasauridae, „Maasechsen“) waren in der gesamten Oberkreide eine weltweit verbreitete Gruppe meeresbewohnender Reptilien.
Sie gehören wie die heute lebenden Schlangen, Echsen und Doppelschleichen zu den Schuppenkriechtieren (Squamata) und erreichten bis zu 18 Meter Länge. Georges Cuvier erkannte Anfang des 19. Jahrhunderts unter anderem am Fossil von Mosasaurus hoffmannii erstmals in der Wissenschaftsgeschichte, dass die Möglichkeit des Aussterbens von Lebewesen besteht.
Merkmale
[Bearbeiten | Quelltext bearbeiten]Die Extremitäten der Mosasaurier hatten sich als Anpassung an die aquatile Lebensweise zu Flossen ausgebildet, welche ähnlich einigen anderen fossilen Meeresreptilien die Kennzeichen des Phänomens der Hyperphalangie aufwiesen (überzählige Fingerglieder). Der lange Schwanz hatte am Ende eine hypocerke Flosse[1], d. h., das Ende der Wirbelsäule biegt sich nach unten und stützt den unteren, größeren Teil der Schwanzflosse. Der langgestreckte Schädel war mit einem sehr kräftigen Kiefer ausgestattet. Im Kiefer von Mosasauriern befanden sich hauptsächlich spitze, meist sehr gleichartige Zähne, doch gab es Ausnahmen wie die Gattung Globidens mit kugelförmigen Brechzähnen. Im mittleren Unterkiefer befand sich ein Gelenk, welches das Öffnen des Maules begünstigte. Mosasaurier hatten viele Wirbel, die sieben Halswirbel stützen zusammen mit den meist 22 Rückenwirbeln und den oft mehr als 100 Schwanzwirbeln den Körper der Mosasaurier. Der Augapfel wird durch einen Ring aus dünnen Knochenplatten um diesen geschützt (Skleralring). Dunkle Spuren (Eumelanin) im fossil erhaltenen Weichgewebe von Mosasaurierfossilien zeigen, dass sie dunkel gefärbt waren und eine helle Bauchseite hatten.[2]
Wie die Ichthyosaurier und Plesiosaurier sollen die Mosasaurier in der Lage gewesen sein, ihre Körpertemperatur auf einem hohen Niveau von 35 bis 39 °C konstant zu halten (Endothermie).[3][4]
Lebensweise
[Bearbeiten | Quelltext bearbeiten]Ernährung
[Bearbeiten | Quelltext bearbeiten]
Das Gebiss der Mosasaurier lässt darauf schließen, dass sich Mosasaurier speziell von großem Nekton wie größeren Fischen ernährten. Fossilfunde, Körperbau und Ausmaße von Mosasauriern lassen darauf schließen, dass Seevögel (damals etwa Hesperornis) und tieffliegende Flugsaurier gefressen wurden, ebenso dürften diverse Meeresreptilien von Mosasauriern gejagt worden sein (wohl hauptsächlich Jungtiere), außerdem Ammoniten. Nach heutigen Erkenntnissen stellten sie der Beute nicht hetzend nach, sondern erlegten sie in schnellem Überraschungsangriff. Einige Mosasaurier wendeten andere Strategien an, um an andere Nahrung zu kommen. So brachen die Vertreter der Gattung Globidens auf Muschelbänken die Muscheln ab und verzehrten sie, ähnlich den sehr viel früher existenten und zu dieser Zeit nur noch fossil erhaltene Placodontiden.
Die Mosasaurier konnten mit den spitzen Zähnen zwar heftige Attacken starten, doch konnte mit diesen Zähnen die Nahrung nicht zerkleinert werden, daher musste die Beute komplett heruntergeschluckt werden. Durch das Gelenk im Unterkiefer konnte der Kiefer sehr weit geöffnet werden und große Nahrung aufnehmen. Gaumenzähne taten ein weiteres.


Fortbewegung
[Bearbeiten | Quelltext bearbeiten]Die großen Extremitäten und die Körperproportionen von Mosasauriern lassen vermuten, dass Mosasaurier ziemlich wendig schwammen. Der Hauptantrieb kam dabei vom kräftigen Ruderschwanz. Bei einer Ausgrabung in Jordanien wurde das Fossil eines Prognathodon gefunden, das eine Schwanzflosse hat, deren längerer Teil nach unten ragt anstatt nach oben wie bei den meisten Haien. Mosasaurier könnten mit dieser Schwanzflosse schneller geschwommen sein als bisher gedacht.[1][5]
Fortpflanzung
[Bearbeiten | Quelltext bearbeiten]Seit den Funden von Mosasaurierembryos innerhalb der Körper von ausgewachsenen Tieren und dem Fund eines großen, weichschaligen Eis in marinen Ablagerungen aus der Oberkreide der Antarktis[6] gilt es als wahrscheinlich, dass Mosasaurier ovovivipar (eilebendgebärend) waren. 1996 beschrieb Gorden Bell die fragmentierten Überreste eines Plioplatecarpus, gefunden in South Dakota, die die Knochen von zwei Embryos bargen. 2001 folgte die Beschreibung des Aigialosauriers Carsosaurus, in dessen hinterem Körperteil vier weit entwickelte Embryos lagen. Die Agialosaurier gelten als Vorfahren der Mosasaurier bzw. als deren Schwestergruppe. Die Lage der Embryos zeigte, dass sie wie Wale und Ichthyosaurier mit dem Schwanz voran geboren wurden. Mosasaurierjunge waren bei der Geburt ein bis zwei Meter lang. In der Niobrara-Formation in Kansas wurde eine Ansammlung von sehr jungen Mosasauriern gefunden, deren fossile Überreste in einem Gebiet abgelagert wurden, das zu ihrer Zeit (Campanium bis unteres Coniacium) mehr als 300 km von der nächsten Küste des Western Interior Seaway entfernt lag. Die Tiere sind also wahrscheinlich auf dem offenen Ozean geboren worden.[7][8]
Forschungsgeschichte und Fossilfunde
[Bearbeiten | Quelltext bearbeiten]Fossile Nachweise von Mosasaurieren sind interkontinental in großer Vielfalt bekannt. Der erste Fund war 1770 in einem Kalksteinbau bei Maastricht. Durch Truppen während der französischen Revolution kam das Fundstück nach Paris, wo Georges Cuvier es als Echse identifizierte. Eine sehr reichhaltige und mit mehr als einem halben Dutzend an Gattungen auch diverse Fauna an Mosasauriern wurde im Ouled-Abdoun-Becken in Marokko entdeckt. Die Funde datieren in das Maastrichtium.[9]
Systematik
[Bearbeiten | Quelltext bearbeiten]Die nächsten Verwandten der Mosasaurier sind die kleinen und noch nicht so sehr an ein marines Leben angepassten Aigialosauridae und Dolichosauridae, mit denen sie zusammen das Taxon Mosasauroidea bilden. Die systematische Stellung der Mosasaurier und ihrer näheren Verwandten innerhalb der Schuppenkriechtiere ist bis heute nicht geklärt und nach wie vor umstritten. Während einige Wissenschaftler in den Mosasauroidea relativ nahe Verwandte der Warane (Varanidae) und der Krustenechsen (Helodermatidae) sehen,[10][11][12] sind andere der Ansicht, dass es sich bei den Mosasauroidea um die Schwestergruppe der Schlangen handelt.[13][14][15][16][17][18][19] Das Taxon, das Schlangen und Mosasaurier vereint, wird Pythonomorpha genannt.
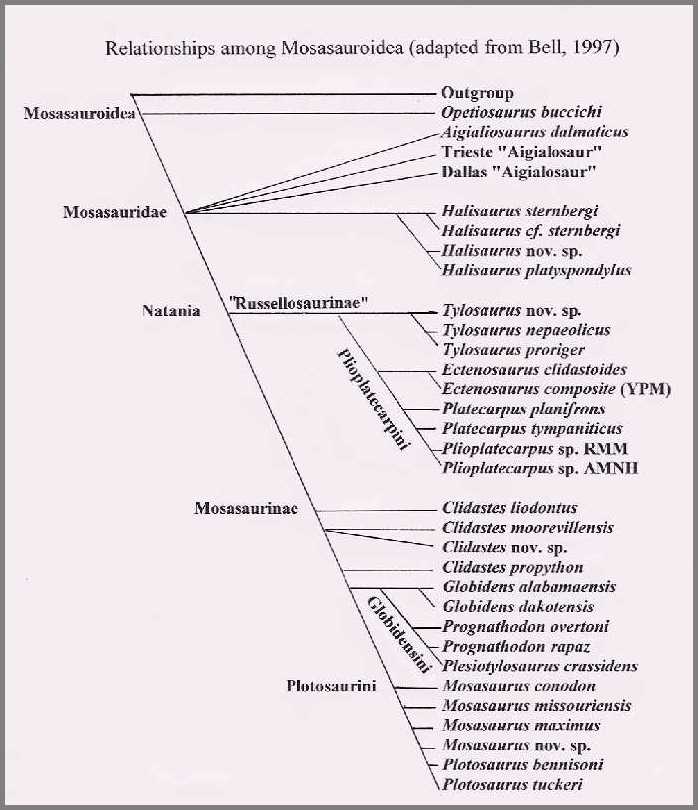
Innerhalb der Mosasaurier werden die primitiveren Halisaurinae und die fortschrittlicheren Natania unterschieden. Innerhalb der Unterfamilie Mosasaurinae finden sich stark abgewandelte Formen, die z. B. ein Brechgebiss entwickelten und, wie Globidens zu Molluskenfressern wurden oder spezialisierte Ichthyosaurierähnliche Fischfresser, wie Plotosaurus. Der krokodilähnliche Goronyosaurus steht isoliert.[20][21]




- Mosasauroidea
- Aigialosaurier (Aigialosauridae)
- Mosasaurier (Mosasauridae)
- Goronyosaurus
- Tethysaurinae[22]
- Halisaurinae
- Natania
- Plesiotylosaurus, Zuordnung umstritten
- Prognathodon, Zuordnung umstritten
- Russelosaurina
- Mosasaurinae
- Amphekepubis
- Eremiasaurus
- Gnathomortis[26]
- Moanasaurus
- Liodon
- Clidastes
- Globidensini
- Plotosaurini
Etymologie
[Bearbeiten | Quelltext bearbeiten]Der französisch-belgisch-niederländische Fluss Maas wird im Lateinischen „Mosa“ genannt. Die ersten Fossilien wurden nahe diesem Fluss gefunden, daher der wissenschaftliche Name Mosasauridae, dies bedeutet also „Maasechse“.
Quellen
[Bearbeiten | Quelltext bearbeiten]Literatur
[Bearbeiten | Quelltext bearbeiten]- Jasper James: Die Kreide. In: Nigel Marven, Jasper James: Monster der Tiefe. Im Reich der Urzeit. vgs, Köln 2004, ISBN 3-8025-1573-0, S. 104–125 (Übersetzung der englischen Originalausgabe von 2003).
- Oskar Kuhn: Reptilien der Urzeit. In: Bernhard Grzimek (Hrsg.): Grzimeks Tierleben. Band 6: Kriechtiere. Unveränderter Nachdruck der dtv-Ausgabe von 1979/80. Bechtermünz, Augsburg 2000, ISBN 3-8289-1603-1, S. 38–72, hier S. 70 f
- Martin Sander: †Mosasauria, Maasechsen. In: Wilfried Westheide, Reinhard Rieger (Hrsg.): Spezielle Zoologie. Band 2: Wirbel- oder Schädeltiere. Gustav Fischer u. a., Stuttgart u. a. 2004, ISBN 3-8274-0900-4, S. 380.
Einzelnachweise
[Bearbeiten | Quelltext bearbeiten]- ↑ a b Johan Lindgren, Hani F. Kaddumi, Michael J. Polcyn: Soft tissue preservation in a fossil marine lizard with a bilobed tail fin. In: Nature Communications. 4, Article number: 2423, September 2013, doi:10.1038/ncomms3423.
- ↑ Johan Lindgren, Peter Sjövall, Ryan M. Carney, Per Uvdal, Johan A. Gren, Gareth Dyke, Bo Pagh Schultz, Matthew D. Shawkey, Kenneth R. Barnes, Michael J. Polcyn: Skin pigmentation provides evidence of convergent melanism in extinct marine reptiles. In: Nature. Bd. 506, Nr. 7489, 2014, S. 484–488, doi:10.1038/nature12899.
- ↑ Aurélien Bernard, Christophe Lecuyer, Peggy Vincent, Romain Amiot, Nathalie Bardet, Éric Buffetaut, Gilles Cuny, François Fourel, François Martineau, Jean-Michel Mazin, Abel Prieur: Regulation of Body Temperature by Some Mesozoic Marine Reptiles. In: Science. Bd. 328, Nr. 5984, 2010, S. 1379–1382, doi:10.1126/science.1187443.
- ↑ T. Lynn Harrell, Alberto Pérez-Huerta, Celina A. Suarez. Endothermic mosasaurs? Possible thermoregulation of Late Cretaceous mosasaurs (Reptilia, Squamata) indicated by stable oxygen isotopes in fossil bioapatite in comparison with coeval marine fish and pelagic seabirds. Palaeontology, 2016; 59 (3): 351, doi:10.1111/pala.12240
- ↑ Nature, 11. September 2013 – MVI/NP: Meeressaurier mit Haifisch-Flosse, Mosasaurier waren wendiger als gedacht – dank haiähnlicher Schwanzflossen in scinexx.de, Datum: 11. September 2013, abgerufen: 11. September 2013
- ↑ Lucas J. Legendre, David Rubilar-Rogers, Grace M. Musser, Sarah N. Davis, Rodrigo A. Otero, Alexander O. Vargas, Julia A. Clarke: A giant soft-shelled egg from the Late Cretaceous of Antarctica. Nature (2020), doi: 10.1038/s41586-020-2377-7
- ↑ Richard Ellis: Sea Dragons. Predators of the Prehistoric Oceans. University Press of Kansas, Lawrence KS 2003, ISBN 0-7006-1269-6, S. 217–218
- ↑ Michael J. Everhart: Rapid evolution, diversification and distribution of mosasaurs (Reptilia; Squamata) prior to the K-T Boundary. In: Tate 2005. 11th annual symposium in paleontology and geology. The Cretaceous-Tertiary boundary. Adaptive radiation after the bottleneck. Tate Geological Museum – Casper College, Casper WY 2005, S. 16–27, online auf oceansofkansas.com.
- ↑ Nathalie Bardet, Alexandra Houssaye, Peggy Vincent, Xabier Pereda Suberbiola, Mbarek Amaghzaz, Essaid Jourani und Saïd Meslouh: Mosasaurids (Squamata) from the Maastrichtian Phosphates of Morocco: Biodiversity, palaeobiogeography and palaeoecology based on tooth morphoguilds. In: Gondwana Research. Bd. 27, 2015, S. 1068–1078, doi:10.1016/j.gr.2014.08.014.
- ↑ Caldwell, M. W., Carroll, R. L. et Kaiser, H. 1995: The pectoral girdle and forelimb of Carsosaurus marchesetti (Aegialosauridae), with a preliminary phylogenetic analysis of Mosasauroids and varanoids. Journal of Vertebrate Paleontology 15(3): 516-531. doi: 10.1080/02724634.1995.10011245
- ↑ Jack L. Conrad: Phylogeny And Systematics Of Squamata (Reptilia) Based On Morphology. In: Bulletin of the American Museum of Natural History. 310, 3. Juni 2008, ISSN 0003-0090, S. 1–182. doi:10.1206/310.1
- ↑ Jack L. Conrad, Jennifer C. Ast, Shaena Montanari, Mark A. Norell: A combined evidence phylogenetic analysis of Anguimorpha (Reptilia: Squamata). In: Cladistics. 27, Nr. 3, 22. Juli 2010, ISSN 0748-3007, S. 230–277. doi:10.1111/j.1096-0031.2010.00330.x
- ↑ Caldwell, M. W. et Lee, M. S. Y. 1997. A snake with legs from the marine Cretaceous of the Middle East. Nature 386:705-709.
- ↑ Caldwell, M. W. 1999. Squamate phylogeny and the relationships of snakes and mosasauroids. Zoological Journal of the Linnean Society 125:115-147. doi: 10.1006/zjls.1997.0144
- ↑ Michael S. Y. Lee: The phylogeny of varanoid lizards and the affinities of snakes. In: Philosophical Transactions of the Royal Society of London B: Biological Sciences. 352, Nr. 1349, 29. Januar 1997, ISSN 0962-8436, S. 53–91. doi:10.1098/rstb.1997.0005
- ↑ Lee, M. S. Y. et Caldwell, M. W. 2000. Adriosaurus and the affinities of mosasaurs, dolichosaurs, and snakes. Journal of Paleontology 74(5):915-937. DOI: 10.1017/S0022336000033102
- ↑ Michael S. Y. Lee: Molecular evidence and marine snake origins. In: Biology Letters. 1, Nr. 2, 22. Juni 2005, ISSN 1744-9561, S. 227–230. doi:10.1098/rsbl.2004.0282
- ↑ Tod W. Reeder; Ted M. Townsend; Daniel G. Mulcahy; Brice P. Noonan; Perry L. Wood, Jr.; Jack W. Sites, Jr.; John J. Wiens (2015). Integrated Analyses Resolve Conflicts over Squamate Reptile Phylogeny and Reveal Unexpected Placements for Fossil Taxa. PLOS ONE. 10 (3): e0118199. doi:10.1371/journal.pone.0118199
- ↑ Tiago R. Simões, Michael W. Caldwell, Mateusz Tałanda, Massimo Bernardi, Alessandro Palci, Oksana Vernygora, Federico Bernardini, Lucia Mancini & Randall L. Nydam: The origin of squamates revealed by a Middle Triassic lizard from the Italian Alps. Mai 2018, Nature 557(7707), DOI: 10.1038/s41586-018-0093-3
- ↑ Kladogramm bei Oceans of Kansas
- ↑ Kladogramm bei Mikko’s Phylogeny Archive
- ↑ László Makádi, Michael W. Caldwell, Attila Ősi: The First Freshwater Mosasauroid (Upper Cretaceous, Hungary) and a New Clade of Basal Mosasauroids. PLOS, December 19, 2012 doi: 10.1371/journal.pone.0051781
- ↑ Takuya Konishi et al. A new halisaurine mosasaur (Squamata: Halisaurinae) from Japan: the first record in the western Pacific realm and the first documented insights into binocular vision in mosasaurs. Journal of Systematic Palaeontology, December 07, 2015; doi:10.1080/14772019.2015.1113447
- ↑ Johan Lindgren, Michael J. Everhart, Michael W. Caldwell: Three-Dimensionally Preserved Integument Reveals Hydrodynamic Adaptations in the Extinct Marine Lizard Ectenosaurus (Reptilia, Mosasauridae). In: PLoS ONE. Bd. 6, Nr. 11, 2011, e27343, doi:10.1371/journal.pone.0027343.
- ↑ Catherine R. C. Strong, Michael W. Caldwell, Takuya Konishi und Alessandro Palci: A new species of longirostrine plioplatecarpine mosasaur (Squamata: Mosasauridae) from the Late Cretaceous of Morocco, with a re-evaluation of the problematic taxon ‘Platecarpus’ ptychodon. Journal of Systematic Palaeontology, 2020, doi:10.1080/14772019.2020.1818322
- ↑ Joshua R. Lively. Redescription and phylogenetic assessment of ‘Prognathodon’ stadtmani: implications for Globidensini monophyly and character homology in Mosasaurinae. Journal of Vertebrate Paleontology, September, 2020, doi: 10.1080/02724634.2020.1784183
- ↑ Nicholas R. Longrich, Nathalie Bardet, Anne S. Schulp und Nour-Eddine Jalil: Xenodens calminechari gen. et sp. nov., a bizarre mosasaurid (Mosasauridae, Squamata) with shark-like cutting teeth from the upper Maastrichtian of Morocco, North Africa. Cretaceous Research, 2021, S. 104764, doi:10.1016/j.cretres.2021.104764
{kind=link}