
 French
French Deutsch
DeutschEquisetaceae , la enciclopedia libre
- Véase también Pteridophyta (plantas vasculares sin semilla)
| Cola de caballo | ||
|---|---|---|
| Rango temporal: Devónico-Holoceno | ||
,_Portland,_Oregon.jpg) | ||
| Taxonomía | ||
| Reino: | Plantae | |
| División: | Monilophyta (antes Pteridophyta) | |
| Clase: | Equisetopsida | |
| Subclase: | Equisetidae | |
| Orden: | Equisetales | |
| Familia: | Equisetaceae (familia nº 4) | |
| Géneros, subgéneros y especies | ||
Equisetum
La clasificación más actualizada hasta género es la de Christenhusz et al. 2011[1][2][3] (basada en Smith et al. 2006,[4] 2008);[5] que también provee una secuencia lineal de las licofitas y monilofitas. | ||
Las equisetáceas (nombre científico Equisetaceae) son una familia monotípica (con un solo género), siendo su género viviente, Equisetum (conocido como cola de caballo), que es el único sobreviviente hasta la actualidad de todas las equisetópsidas, las cuales tienen un extenso registro fósil entre el Devónico y el Carbonífero. Las equisetáceas son morfológicamente muy distintivas, con un tallo con crestas y valles, hojas reducidas dispuestas en verticilos, y hojas fértiles transformadas en esporangióforos unidas en estróbilos terminales. Este género consta de unas 18 especies agrupadas en tres subgéneros según la Plants of the World online (POWO). Además, las especies dentro de cada subgénero hibridan con facilidad.
Caracteres
[editar] Referencias: n : generación haploide, 2n : generación diploide, m! : mitosis, M! : meiosis, F! : fecundación |
Los equisetos son plantas vasculares con ciclo de vida haplodiplonte donde la alternancia de generaciones es bien manifiesta, con esporófito y gametófito multicelulares e independientes, con esporas como unidad de dispersión y de resistencia. El gametófito es un "talo" (cuerpo sin organización), y el esporófito es un "cormo" (con raíz, vástago y sistema vascular). Debido a estas características tradicionalmente se las agrupa con las "pteridofitas".
Los tallos, fotosintéticos, crecen a partir de un rizoma subterráneo mediante un meristema que consta de una célula apical piramidal que se divide en tres direcciones.
Los tallos presentan nudos y entrenudos bien diferenciados, se dice que los nudos son "ensanchados" ("swollen", "joint-like"). La superficie del tallo en los estrenudos es acanalada: presenta crestas y valles longitudinales (pliegues y estrías, carenas y valéculas), carácter único en monilofitas. Las células de la epidermis del tallo depositan sílice en la superficie del tallo que actúa como refuerzo de sostén. El tallo es usualmente hueco, de forma similar a una caña; observándose en el corte transversal del tallo un hueco central y otros huecos pequeñitos del lado interno de las crestas y los valles.
Las hojas (eufilos reducidos) son muy pequeñas (menos de 2 cm de largo usualmente) y tienen aspecto escamoso, se disponen alrededor del tallo de forma verticilada (más de 2 hojas en el mismo nudo), y son connadas (están soldadas entre sí por su base), formando una vaina o corona —"sheath"— que a veces es más o menos engrosada. Las hojas solo al principio son fotosintéticas secándose luego [cita requerida]. Solo son irrigadas por un haz vascular no ramificado.[cita requerida]
-
 Detalle de tallo y un verticilo de hojas de Equisetum hyemale Se observan las crestas y valles, y las hojas connadas en la base formando la vaina o corona.
Detalle de tallo y un verticilo de hojas de Equisetum hyemale Se observan las crestas y valles, y las hojas connadas en la base formando la vaina o corona. -
 Detalle de tallo y un verticilo de hojas de Equisetum arvense Se observan las crestas y valles, y las hojas connadas en la base formando la vaina o corona.
Detalle de tallo y un verticilo de hojas de Equisetum arvense Se observan las crestas y valles, y las hojas connadas en la base formando la vaina o corona. -
 Detalle de tallo y dos verticilos de hojas de Equisetum fluviatile Se observan las crestas y valles, y las hojas connadas en la base formando la vaina o corona.
Detalle de tallo y dos verticilos de hojas de Equisetum fluviatile Se observan las crestas y valles, y las hojas connadas en la base formando la vaina o corona.



De los nudos pueden partir una gran cantidad de ramas en el subgénero Equisetum, de disposición verticilada, y de morfología similar a la del tallo.
-
 Detalle de nudo de Equisetum palustre Nacimiento de las ramas.
Detalle de nudo de Equisetum palustre Nacimiento de las ramas. -
 Esporófito joven de Equisetum telmateia Una especie con ramas.
Esporófito joven de Equisetum telmateia Una especie con ramas. -
 Hábito de Equisetum arvense Se observan las ramas.
Hábito de Equisetum arvense Se observan las ramas.



Al final de las ramas fértiles se encuentra el estróbilo, formado por los verticilos de hojas fértiles (que en este clado se llaman "esporangióforos").
El eje del estróbilo puede ser verde o no verde, y nunca es ramificado. Los esporangióforos son peltados (con pie), no fotosintéticos. Los esporangióforos jóvenes tienen forma de pulvínulo y luego toman forma hexagonal (en vista superficial), debido a la fuerza mecánica que ejercen entre ellos.
-
 Aspecto general de estróbilo de Equisetum hyemale
Aspecto general de estróbilo de Equisetum hyemale -
 Estróbilo joven de Equisetum arvense Se observan los esporangios colgando hacia adentro de los esporangióforos peltados. Planta preservada en líquido.
Estróbilo joven de Equisetum arvense Se observan los esporangios colgando hacia adentro de los esporangióforos peltados. Planta preservada en líquido. -
 Estróbilo seco de Equisetum arvense
Estróbilo seco de Equisetum arvense



En la cara interna del esporangióforo se originan los esporangios, que penden por debajo del ápice expandido del esporangióforo.
-
 Detalle de estróbilo de Equisetum Se observan los esporangióforos. Planta preservada en líquido.
Detalle de estróbilo de Equisetum Se observan los esporangióforos. Planta preservada en líquido. -
 Corte transversal de estróbilo de Equisetum Se observan los esporangios colgando hacia adentro de los esporangióforos peltados. Planta preservada en líquido.
Corte transversal de estróbilo de Equisetum Se observan los esporangios colgando hacia adentro de los esporangióforos peltados. Planta preservada en líquido. -
 Corte longitudinal de estróbilo de Equisetum arvense Se observan los esporangios colgando hacia adentro de los esporangióforos peltados.
Corte longitudinal de estróbilo de Equisetum arvense Se observan los esporangios colgando hacia adentro de los esporangióforos peltados.



Los esporangios presentan un engrosamiento secundario de la pared de forma helicoide (Bateman 1991). Los esporangios son de tipo eusporangiado: grandes, sin anillo, elongados, producen muchas (más de 1000) esporas por esporangio.
Los estróbilos se alargan al madurar para separar los esporángióforos y permitir la dispersión de las esporas (cita requerida).
Las esporas son dispersadas por el viento.
Las esporas son esféricas, verdes, con apertura circular, y con cuatro a seis eláteres (expansiones largas en forma de correas) enrollados alrededor de la espora, que rápidamente se enderezan y asisten al movimiento de la espora cuando sale del esporangio en la dehiscencia.
-
 Foto al microscopio de esporas mojadas de Equisetum Los eláteres están enrollados alrededor de las esporas.
Foto al microscopio de esporas mojadas de Equisetum Los eláteres están enrollados alrededor de las esporas. -
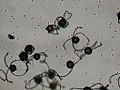 Foto al microscopio de esporas secas de Equisetum Los eláteres erectos están preparados para ayudar a la espora a dispersarse transportada por el viento.
Foto al microscopio de esporas secas de Equisetum Los eláteres erectos están preparados para ayudar a la espora a dispersarse transportada por el viento.


Las equisetáceas son plantas homospóricas (todas las esporas son iguales). En las plantas homospóricas en general, las esporas poseen la capacidad de producir gametófitos capaces de originar gametos masculinos y femeninos (las esporas producen gametófitos hermafroditas). Sin embargo existe una controversia acerca de si los gametófitos de las equisetáceas son hermafroditas o no. Algunos gametófitos producen solo anteridios (que dan gametos masculinos), otros gametófitos producen solo arquegonios (que dan gametos femeninos). Según Judd et al. (2002), al menos los gametófitos "femeninos" se vuelven bisexuales más tarde.
Los gametofitos son verdes, superficiales.
Los anteridios producen una gran cantidad de espermatozoides multiflagelados. La morfología de los espermatozoides vincula claramente a las equisetáceas con las demás monilofitas.
Número de cromosomas x = 108.
Sistemática
[editar]- Introducción teórica en Filogenia

Tanto la morfología como los análisis moleculares de ADN (hechos sobre las secuencias plastídicas rbcL, trnL-F, rps4) indican que es monofilético (Pryer et al. 2001[6] y 2004,[7] Des Marais et al. 2003,[8] Guillon 2004).[9]
Este género está compuesto por dos subgéneros, la hibridación es frecuente entre especies del mismo subgénero pero no entre especies de subgéneros diferentes. Los análisis moleculares de ADN hechos sobre las 15 especies (basados en un análisis combinado de dos marcadores en el cloroplasto, trnL-F y rbcL), demuestran un robusto consenso para la monofilia de los dos subgéneros. Un solo resultado es una concepción nueva: la especie sudamericana, Equisetum bogotense, no pertenece a ninguno de los dos subgéneros sino más bien es hermana de las otras 14 especies.
El análisis combinado de morfología y secuencias de ADN (sobre las secuencias cloroplastídicas rbcL, atpB, rps4, y sobre la secuencia nuclear para la subunidad pequeña de ADNr) lo ubicaron como un clado hermano de las marattiáceas y de los helechos leptosporangiados, formando entre los tres un clado hermano de Ophioglossaceae y Psilotaceae (Pryer et al. 2001[6] y 2004[7]). Los caracteres morfológicos que incluyen a Equisetum con el resto de los helechos en el clado Monilophyta, son la morfología de los espermatozoides (Renzaglia et al. 2000)[10] y caracteres de las raíces (Kato 1983).[11] El cladograma resultante es el que muestra el siguiente gráfico:
 |
Taxonomía
[editar]- Introducción teórica en Taxonomía
La clasificación más actualizada hasta género es la de Christenhusz et al. 2011[1][2][3] (basada en Smith et al. 2006,[4] 2008);[5] que también provee una secuencia lineal de las licofitas y monilofitas.
- Subclase: II Equisetidae Warm., Osnov. Bot.: 221 (1883).
- Orden D. Equisetales DC. ex Bercht. & J.Presl, Přir. Rostlin: 271 (1820).
- 1 familia.
- Familia 4. Equisetaceae Michx. ex DC., Essai Propr. Méd. Pl.: 49 (1804).
- 1 género (Equisetum). Referencias: Des Marais et al. (2003), Guillon (2004, 2007), Hauke (1963, 1978), Schaffner (1930).
- Nota: La ubicación de Equisetidae todavía es poco clara. Algunos estudios basados en ADN plastídico lo ubican como hermano de Marattiidae (Pryer et al. 2004, Smith et al. 2006a), pero Rai & Graham (2010) encontraron que Equisetidae es hermano del resto de los helechos. Esta relación es más consistente con el récord fósil (Taylor et al. 2009) y está de acuerdo con la morfología del grupo, si bien pensamos que debería ser incluido en el linaje de helechos es decir Polypodiopsida (en el sentido de Pryer et al. 2001).
Clasificación sensu Smith et al. 2006
[editar]Clados y taxones superiores: Plantae (clado), Viridiplantae, Streptophyta, Streptophytina, Embryophyta, Tracheophyta, Euphyllophyta, Monilophyta, Clase Equisetopsida, Orden Equisetales, familia Equisetaceae, género Equisetum.
Equisetum proviene del latín equus (caballo) y seta (cerda).
Nombre vulgar: cola de caballo.
Circunscripción: tres subgéneros:
- Especie sudamericana.
- Subgénero Paramochaete. 1 especie.
- Estomas superficiales y tallos ramificados:
- Subgénero Equisetum. 8 especies.
- Estomas hundidos y tallos no ramificados.
- Subgénero Hippochaete. 9 especies.
- Equisetum giganteum
- Equisetum hyemale L.
- Equisetum laevigatum
- Equisetum myriochaetum Schltr. & Cham.
- Equisetum praealtum Raf
- Equisetum ramosissimum Desf.
- Equisetum scirpoides Michx.
- Equisetum variegatum Schleich.
- Equisetum xylochaetum Mett.
Otras clasificaciones
[editar]En las clasificaciones más tradicionales como el Engler, al tener las hojas reducidas, parecidas a microfilos, el clado fue pensado como una pteridofita primitiva, y fue asignado a su propia clase, considerado más reciente que los psilotos (que tienen las hojas aún más reducidas), y más antiguo que los demás helechos.
Con tantos caracteres inequívocos (la morfología del tallo, las hojas verticiladas, los esporangióforos) el clado fue rápidamente reconocido como tal, por lo que la circunscripción (es decir, de qué subtaxones está compuesto) no ha variado. La clasificación sensu Engler es la siguiente:
- Reino Plantae (polifilético), división Embryophyta asiphonogama (parafilético), subdivisión Pteridophyta (parafilético), clase Sphenopsida (monofilético, equivalente a Equisetopsida sensu Smith et al.), orden Equisetales, familia Equisetaceae (monofilético, equivalente a Equisetaceae sensu Smith et al.).
Ecología
[editar]Ampliamente distribuidas: se encuentran en todo el mundo excepto en Australia, Nueva Zelanda y Antártida. El mayor número de especies se encuentra entre los 40° y los 60° de latitud norte.
Plantas herbáceas, rizomatosas y perennes. Morfología bambusoide (no emparentadas con los bambúes). Tallos normalmente de menos de 1 metro de altura, pero pueden llegar a los 8 metros en E. giganteum. Tallos de 2-4 cm de diámetro.
Terrestres a acuáticas. Estas plantas son colonizadores de áreas deforestadas, márgenes de lagos, y humedales.
Evolución
[editar]Los parientes fósiles de Equisetum aparecen desde el Devónico (hace unos 408-360 millones de años) y luego se han vuelto más abundantes y relativamente pequeños (menos de 1 m de altura) adaptados al sotobosque de los bosques del Carbonífero.
Algunos miembros del clado de las equisetópsidas tienen tallos que llegaron hasta los 20 metros de altura, pero se extinguieron en el Carbonífero tal como las lycophytas gigantes de ese tiempo.
Los primeros fósiles claramente asignables al lineaje de las equisetáceas (claros ancestros de Equisetum) son del Eoceno (hace unos 54-38 millones de años), pero el género puede extenderse hasta el Pérmico, hace más de 300 millones de años.
Importancia económica
[editar]La sílice presente en los tallos los hace buenos para restregar, los colonos de Norteamérica los usaban para fregar los utensilios de cocina, una práctica que aparentemente dio origen a otro nombre vulgar de esta planta, "junco para refregar" ("scouring rush"). Los artesanos tradicionales del dorado artístico usan ya desde la Edad Media los cálamos secos de esta planta para pulir las superficies doradas, por ejemplo, imágenes escultóricas policromadas y marcos para lienzos, y conseguir así contrastes de brillo y mate.
Uso medicinal
[editar]El Equisetum giganteum L. y otras equisetáceas son ampliamente utilizadas como plantas medicinales utilizándose su parte aérea como diurético. En Argentina Equisetum giganteum es una planta medicinal oficial ya que posee una monografía en la Farmacopea Nacional Argentina VI Edición.
Véase también
[editar]- Clasificación de los organismos vegetales.
- Para ver una lista completa de los taxones ver Sistema de clasificación de monilophytas de Smith 2006.
- Para una descripción básica de morfología y anatomía de los helechos ver Pteridophyta.
Referencias
[editar]- A. R. Smith, K. M. Pryer, E. Schuettpelz, P. Korall, H. Schneider, P. G. Wolf. 2006. "A classification for extant ferns". Taxon 55(3), 705-731 (pdf aquí)
- Judd, W. S. Campbell, C. S. Kellogg, E. A. Stevens, P.F. Donoghue, M. J. 2002. Plant systematics: a phylogenetic approach, Second Edition. Sinauer Axxoc, USA.
- Stevens, P. F. (2001 en adelante). Angiosperm Phylogeny Website. Versión 7 (mayo de 2006) y actualizado regularmente.
Referencias citadas
[editar]- ↑ a b Christenhusz et al. 2011. A linear sequence of extant families and genera of lycophytes and ferns. Phytotaxa 19: 7-54. (pdf)
- ↑ a b Preface to “Linear sequence, classification, synonymy, and bibliography of vascular plants: Lycophytes, ferns, gymnosperms and angiosperms” pdf
- ↑ a b Corrections to Phytotaxa 19: Linear sequence of lycophytes and ferns pdf
- ↑ a b A. R. Smith, K. M. Pryer, E. Schuettpelz, P. Korall, H. Schneider, P. G. Wolf. 2006. "A classification for extant ferns". Taxon 55(3), 705-731 ( pdf)
- ↑ a b Smith, A.R., Pryer, K.M., Schuettpelz, E., Korall, P., Schneider, H., & Wolf, P.G. (2008) Fern classification, pp. 417–467 en: Ranker, T.A., & Haufler, C.H. (eds.), Biology and Evolution of Ferns and Lycophytes. Cambridge , Cambridge University Press.
- ↑ a b Pryer, Kathleen M., Harald Schneider, Alan R. Smith, Raymond Cranfill, Paul G. Wolf, Jeffrey S. Hunt y Sedonia D. Sipes. 2001. "Horsetails and ferns are a monophyletic group and the closest living relatives to seed plants". Nature 409: 618-622 (pdf aquí).
- ↑ a b Pryer, Kathleen M., Eric Schuettpelz, Paul G. Wolf, Harald Schneider, Alan R. Smith y Raymond Cranfill. 2004. "Phylogeny and evolution of ferns (monilophytes) with a focus on the early leptosporangiate divergences". American Journal of Botany 91:1582-1598 (pdf aquí).
- ↑ Des Marais, D. L., Smith, A. R., Britton, D. M., y Pryer, K. M. 2003. Phylogenetic relationships and evolution of extant horsetails, Equisetum, based on chloroplast DNA sequence data (rbcL and trnL-F). Int. J. Pl. Sci. 164: 737-751. (pdf aquí)
- ↑ Guillon, J. M. 2004. Phylogeny of horsetails (Equisetum) based on the chloroplast rps4 gene and adjacent noncoding sequences. Syst. Bot. 29: 251-259. (resumen aquí)
- ↑ Renzaglia, K. S., Duff, R. J., Nickrent, D. L., y Garbary, D. J. 2000. Vegetative and reproductive innovations of early land plants; implications for an unified phylogeny. Philos. Trans. Ser. B. 355: 769-793.
- ↑ Kato, M. 1983. The classification of major groups of pteridophytes. J. Fac. Sci. Univ. Tokyo, Sect. 3, Bot. 13: 263-283.
Enlaces externos
[editar] Wikimedia Commons alberga una categoría multimedia sobre Equisetaceae.
Wikimedia Commons alberga una categoría multimedia sobre Equisetaceae. Wikispecies tiene un artículo sobre Equisetaceae.
Wikispecies tiene un artículo sobre Equisetaceae.
| Control de autoridades |
|
|---|
 Datos: Q517643
Datos: Q517643- Multimedia: Equisetaceae / Q517643
- Especies: Equisetaceae
j