
 French
French Deutsch
DeutschBasidiomyceten
| Basidiomycota Fossiel voorkomen: Vroeg-Carboon[1] – heden | |||
|---|---|---|---|
 | |||
| Diversiteit van de basidiomyceten | |||
| Taxonomische indeling | |||
| |||
| Stam | |||
| Basidiomycota R.T.Moore (1980) | |||
| Afbeeldingen op | |||
| Basidiomycota op | |||
| |||
Basidiomyceten (Basidiomycota) vormen een stam binnen het rijk van de schimmels (Fungi). Basidiomyceten worden ook wel steeltjeszwammen[2] genoemd. Kenmerkend voor deze stam is dat de sporen in een basidium (steeltje) gevormd worden. Deze steeltjes zijn enkel microscopisch waarneembaar. Meestal zijn ze met elkaar vergroeid tot een kiemvlies of hymenium. In het kiemvlies komen vaak cystiden voor. Dat zijn steriele cellen die een rol spelen bij de dosering van sporenuitstoot tijdens het sporuleren.
Vrijwel alle basidiomyceten zijn filamenteuze schimmels die zich geslachtelijk voortplanten via de vorming van gespecialiseerde sporen, de zogenaamde basidiosporen. Sommige soorten kunnen zich uitsluitend ongeslachtelijk voortplanten. Deze soorten zijn te herkennen door morfologische overeenkomsten, de vorming van een onderscheidend anatomisch kenmerk (de gesp-verbinding), celwandcomponenten en definitief door fylogenetische moleculaire analyse.
Voortplanting
[bewerken | brontekst bewerken]
a= aecidium; sp= spermogonium;
ur= uredospore; t= teleutospore
In het algemeen komt zowel 'heterothallie' als 'homothallie' voor bij de basidiomyceten. Heterothallie komt bij de basidiomyceten het meeste voor. Ongeveer 90% van de Agaricomycotina zijn heterothallisch.[3]
Sommige soorten hebben van alle schimmels de meest complexe seksuele voortplanting. Zo heeft zwarte roest (Puccinia graminis) een monocotyle waardplant voor de vegetatieve fase en een dicotyle waardplant voor de generatieve fase. De schimmel vormt vijf verschillende sporen: uredosporen, teleutosporen, basidiosporen, spermatiën en aecidiosporen.
De paringscompatibiliteit wordt bij de basidiomyceten in twee paringstypen ingedeeld: tetrapolair en bipolair.
- Een bipolaire schimmel heeft één gen voor paring met de allelen A of a.
- Een tetrapolaire schimmel heeft twee, onafhankelijke genen voor paring. Ze worden met A en B (bij de onderstam Agaricomycotina) of a en b (bij de onderstammen Ustilaginomycotina en Pucciniomycotina) aangeduid. Het a gen heeft twee allelen en het b gen meer dan twee. De twee basidiosporen (sporidia) moeten verschillende allelen van beide genen hebben willen ze zich met elkaar verenigen. Zo is een basidiospore met a1b2 verenigbaar met een basidiospore met a2b1 allelen. Het a gen codeert voor een feromoon, waardoor de vorming van een conjugatiebuis en versmelting in gang gezet wordt. Het b gen codeert voor regulerende eiwitten die zich aan het DNA-binden. Er kunnen meer dan twee allelen per gen voorkomen. Builenbrand heeft meer dan 25 b, maar slechts 2 a allelen.[4] Coprinopsis cinerea heeft meer dan 240 allelen van zowel A als B en Schizophyllum commune heeft meer dan 339 allelen van A en 64 van B.[5]
Alleen schimmels met verschillende allelen zijn compatibel en kunnen paren. De paring begint met de vorming van een conjugatiebuis en de uitwisseling van celkernen. Er ontstaan dikaryotische (tweekernige) hyfen met de afzonderlijke haploïde kernen van beide ouders. Onder de juiste milieuomstandigheden wordt er een vruchtlichaam met basidia op een hymenofoor gevormd. De hymenofoor is de drager van de basidia op het hymenium. Een basidium ontstaat meestal door gespvorming. Daarnaast zijn er ook soorten die geen gespvorming hebben. Hier ontstaat een basidium door knopvorming van de eindcel aan een dikaryotische hyfe. Een derde variant is de vorming van een probasidium.
Op het basidium vindt de vorming van basidiosporen (sporidia) plaats via karyogamie en meiose. De dikaryotische hyfen behouden vaak de gesp-verbinding. De vorming van deze verbindingen wordt gereguleerd door de beide paringsgenen. Voorbeelden zijn de branden, zoals builenbrand (Ustilago maydis).[6][7] en de paddenstoel waaiertje (Schizophyllum commune).[8]
-
 Paring van twee compatibele monokaryons
Paring van twee compatibele monokaryons -
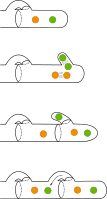 gespvorming
gespvorming -




De generatieve fase van de roesten vindt vaak plaats op een tussenwaardplant. Bij zwarte roest is dit de zuurbes (Berberis vulgaris). Hier vindt de generatieve fase plaats, waarbij spermatiën en aecidiosporen gevormd worden. De teleutospore is de dikwandige, gekleurde rustspore, die in de loop van het seizoen gevormd wordt in het telium (vruchtlichaam) op de andere waardplant. Het telium wordt net voor het afsterven van deze waardplant gevormd. De gesteelde teleutospore is tweecellig en heterothallisch. In het voorjaar kiemt de teleutospore en vormt een basidium, waarop de basidiosporen gevormd worden. De basidiospore kiemt op berberis en vormt daar een spermogonium met spermatiën en receptieve hyfen. Na bevruchting van een receptieve hyfe van het spermogonium door een spermatium met een ander paringstype wordt een aecidium met haploïde-dikaryotische aecidiosporen gevormd. Ook kan er een aecidium gevormd worden uit twee paringshyfen met verschillend paringstype. De aecidiospore kiemt op de graanplant, waarna een uredinium gevormd wordt. Het uredinium vormt uredosporen, die vervolgens weer een uredinium vormen. Later in het seizoen vormt ten slotte een uredinium teleutosporen, doordat het overgaat in een telium of vormt het mycelium een telium.
-
 Levenscyclus Agaricus-soorten, zoals van de champignon. P=paring, K=karyogamie, M=meiose.
Levenscyclus Agaricus-soorten, zoals van de champignon. P=paring, K=karyogamie, M=meiose. -
 Levenscyclus zwarte roest
Levenscyclus zwarte roest
_nl_txt.png)

Aantasting van stamboon en pronkboon door roest (Uromyces appendiculatus var. appendiculatus).
-
 Kiembuis uredospore met appressorium
Kiembuis uredospore met appressorium -
 Kiembuizen uredosporen
Kiembuizen uredosporen -
 Uredosporen (licht gekleurd) en teleutosporen (donkergekleurd)
Uredosporen (licht gekleurd) en teleutosporen (donkergekleurd) -
 Telia
Telia -
 Spermogonia (links) en Aecidia (rechts)
Spermogonia (links) en Aecidia (rechts) -
 Spermogonia
Spermogonia






Taxonomische indeling
[bewerken | brontekst bewerken]Een 2016 indeling geven Holt & Iudica:[9]
Oudere indeling
[bewerken | brontekst bewerken]De taxonomische indeling van de stam Basidiomycota is volgens de Index Fungorum (op 9-9-2008)[10] als volgt:
Daarnaast, ongeplaatst (incertae sedis), de orde Malasseziales plus enkele geslachten. | Eerder[11] was de indeling:
|
Zie ook
[bewerken | brontekst bewerken]Externe links
[bewerken | brontekst bewerken]- nomenclatorische gegevens van de Basidiomycota op de Index Fungorum
- Indeling van de Basidiomycota op de Index Fungorum
- ↑ (en) Krings M, Dotzler N, Galtier J, Taylor TM. (2011). Oldest fossil basidiomycete clamp connections. Mycoscience 52 (1): 18-23. DOI: 10.1007/S10267-010-0065-4Get.
- ↑ Taxoninfo in het Nederlands Soortenregister
- ↑ James, Timothy, Sex in Fungi: Molecular Determination and Evolutionary Implications, 2007, ASM Press, Washington DC, 317–331, J. Heitman, J. W. Kronstad, J. W. Taylor and L. A. Casselton, Analysis of mating type locus organization and synteny in mushroom fungi: Beyond model species
- ↑ Hsueh, Y. P., Heitman, J., Orchestration of sexual reproduction and virulence by the fungal mating-type locus, Current Opinion in Microbiology, 2008, 11, 6, 517–524, doi:10.1016/j.mib.2008.09.014
- ↑ Casselton, L. A., Kües, U., J. Heitman, Sex in Fungi: Molecular Determination and Evolutionary Implications, 2007, ASM Press, Washington, DC, 283–300, editors: J. W. Kronstad, J. W. Taylor, L. A. Casselton, The Origin of Multiple Mating Types in the Model Mushrooms Coprinopsis cinerea and Schizophyllum commune
- ↑ Bakkeren, G, Kronstad, J. W., The Pheromone Cell Signaling Components of the Ustilago a Mating-Type Loci Determine Intercompatibility Between Species, Genetics, 1996, 143, 1601–1613
- ↑ Schulz, B, Banuett, F Dahl, M Schlesinger, R Schäfer, W Martin, T Herskowitz, I Kahmann, R, The b alleles of U. maydis, whose combinations program pathogenic development, code for polypeptides containing a homeodomain-related motif, Cell, 1990, 60, 2, 295–306, doi:10.1016/0092-8674(90)90744-y
- ↑ Raudaskoski, M, Kothe, E, Basidiomycete mating type genes and pheromone signaling, Eukaryotic, Cell, 2010, 9, 6, 847–859, doi:10.1128/ec.00319-09
- ↑ Holt, J.R. & C.A. Iudica (2016) Diversity of Life. Subkingdom Dikarya'
- ↑ Index Fungorum Indeling van de stam Blastocladiomycota
- ↑ 21 oktober 2004; conform de Dictionary of the Fungi, 9th Edition, 2001