
 French
French Deutsch
DeutschКавеолы — Википедия
{kind=link}
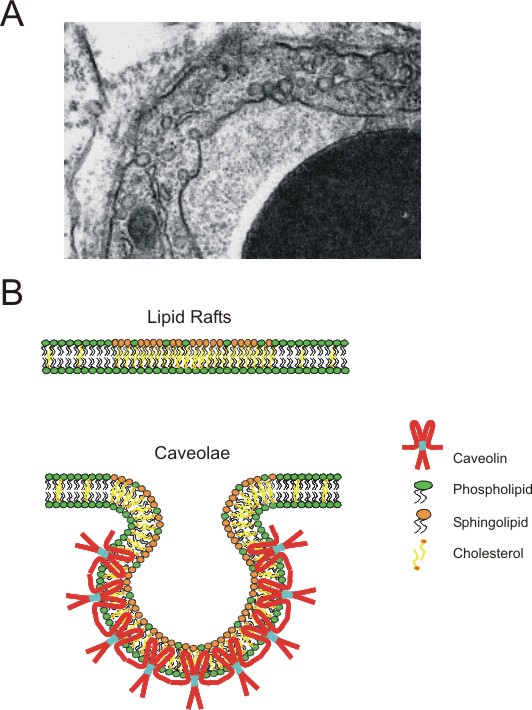
Кавео́лы (от лат. caveola — «малая пещера») — небольшие (размером 50—100 нм) колбообразные впячивания плазматической мембраны в клетках позвоночных многих типов, в особенности эндотелиальных клетках (где они и были впервые обнаружены[1]), адипоцитах и альвеолоцитах I типа (кавеолы могут составлять 30—70 % мембран этих клеток)[2]. В состав кавеол входит ключевой белок — кавеолин[3], а также такие липиды, как холестерин и сфинголипиды. Кавеолы участвуют в передаче клеточных сигналов[4], эндоцитозе, онкогенезе, заражении клетки рядом патогенных бактерий и вирусов[5][6][7].
Строение
[править | править код]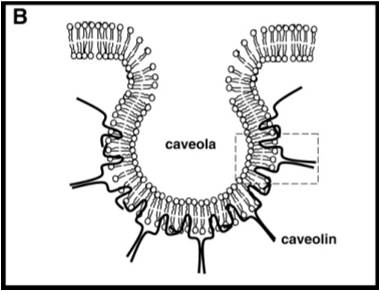{kind=link}
{kind=link}
Образование и поддержание структуры кавеол обеспечивается в основном белком кавеолином массой 21 кДа. У млекопитающих существуют три гена, кодирующих кавеолины: Cav1[англ.], Cav2[англ.] и Cav3[англ.][8]. Эти белки имеют общую топологию: они представляют собой интегральные мембранные белки с двумя глобулярными доменами, соединёнными шпилькообразным гидрофобным доменом, который связывает белок с цитоплазматической стороной плазматической мембраны. Дополнительно белок заякоривают в мембране три пальмитоильные группы, связанные с С-концевым глобулярным доменом. Кавеолин связывается с холестерином в мембране, вызывая образование впячивания мембранного липидного бислоя — кавеолы. Кавеолы — это необычные липидные рафты: они содержат оба липидных слоя мембраны, и цитоплазматический, с которым связывается кавеолин, и обращённый наружу, который является типичным сфинголипид-холестероловым рафтом со связанными посредством GPI-якоря белками[3].
Функции
[править | править код]Кавеолы участвуют во множестве клеточных процессов, среди которых мембранный транспорт и формирование клеточного ответа на внешний сигнал[3]. Одной из важнейших функций кавеол является их участие в эндоцитозе. В отличие от клатриновых, COPI[англ.]- и COPII[англ.]-везикул, инвагинация кавеол и захват груза обусловлены липидным составом мембраны, а не сборкой цитоплазматической белковой оболочки. Может быть, кавеолины принимают участие в стабилизации рафтов с определёнными мембранными белками. Отшнуровывание кавеол от плазматической мембраны опосредовано GTPазой динамином II; после этого кавеолы доставляют своё содержимое либо в кавеосому — особый компартмент, похожий на эндосому, либо в ходе трансцитоза переносят его на противоположный конец полярной клетки. Кавеосома может далее слиться с другой кавеосомой или с первичной эндосомой. С помощью такого эндоцитоза происходит, например, трансцитоз альбумина эндотелиальными клетками и интернализация инсулиновых рецепторов у адипоцитов. Стоит отметить, что, так как кавеолины — это интегральные белки мембраны, они не диссоциируют от везикул после эндоцитоза, а вместо этого доставляются в компартменты-мишени, где существуют как мембранные белки[1]. Кроме того, в кавеосомах сохраняется нейтральный рН и отсутствуют маркеры, характерные для ранних эндосом[8][9].
Рецепторы инсулина и других факторов роста, а также некоторых GTP-связывающих белков и протеинкиназ, связаны с трансмембранной передачей сигнала, которая, по-видимому, локализуется в рафтах и, возможно, кавеолах[3]. Показано, что кавеолины могут связывать некоторые сигнальные молекулы и таким образом регулировать передачу сигнала. Кавеолы также вовлечены в регуляцию мембранных каналов[англ.] и кальциевые сигнальные пути[англ.][9].
Кавеолы могут быть использованы некоторыми патогенами для проникновения в клетку, и таким образом они избегают деградации в лизосомах. Впрочем, некоторые бактерии используют не просто единичные кавеолы, а богатые кавеолами участки мембран. К патогенам, использующим такой путь, относятся вирус SV40, полиомавирус, а также некоторых бактерии, в числе которых — некоторые штаммы Escherichia coli, Pseudomonas aeruginosa и Porphyromonas gingivalis[англ.][9].
Кавеолы могут быть вовлечены в регуляцию липидного обмена. В адипоцитах наблюдается высокий уровень экспрессии кавеолина Cav1. Кавеолин связывается с холестерином, жирными кислотами и жировыми каплями, тем самым принимая участие в управлении липидным обменом[9].
Наконец, кавеолы могут функционировать как механосенсоры[англ.] в клетках некоторых типов. В частности, в клетках эндотелия они задействованы в ощущении скорости тока крови по сосудам. В гладкой мускулатуре кавеолин Cav1 участвует в формировании чувства растяжения мышц[9].
Ингибиторы
[править | править код]Известны некоторые ингибиторы работы кавеол, среди которых филипин III, генистеин и нистатин[8].
См. также
[править | править код]Примечания
[править | править код]- ↑ 1 2 Альбертс и др., 2013, с. 1213.
- ↑ Dr. Babak Razani, Dr. Michael Lisanti. The Role of Caveolae and the Caveolins in Mammalian Physiology. // Reviews in Undergraduate Research. — № 1. Архивировано 24 сентября 2015 года.
- ↑ 1 2 3 4 Nelson, Cox, 2008, p. 386.
- ↑ Anderson R. G. The caveolae membrane system. (англ.) // Annual review of biochemistry. — 1998. — Vol. 67. — P. 199—225. — doi:10.1146/annurev.biochem.67.1.199. — PMID 9759488.
- ↑ Frank P. G., Lisanti M. P. Caveolin-1 and caveolae in atherosclerosis: differential roles in fatty streak formation and neointimal hyperplasia. (англ.) // Current opinion in lipidology. — 2004. — Vol. 15, no. 5. — P. 523—529. — PMID 15361787.
- ↑ Li X. A., Everson W. V., Smart E. J. Caveolae, lipid rafts, and vascular disease. (англ.) // Trends in cardiovascular medicine. — 2005. — Vol. 15, no. 3. — P. 92—96. — doi:10.1016/j.tcm.2005.04.001. — PMID 16039968.
- ↑ Pelkmans L. Secrets of caveolae- and lipid raft-mediated endocytosis revealed by mammalian viruses. (англ.) // Biochimica et biophysica acta. — 2005. — Vol. 1746, no. 3. — P. 295—304. — doi:10.1016/j.bbamcr.2005.06.009. — PMID 16126288.
- ↑ 1 2 3 Lajoie P., Nabi I. R. Lipid rafts, caveolae, and their endocytosis. (англ.) // International review of cell and molecular biology. — 2010. — Vol. 282. — P. 135—163. — doi:10.1016/S1937-6448(10)82003-9. — PMID 20630468.
- ↑ 1 2 3 4 5 Parton R. G., Simons K. The multiple faces of caveolae. (англ.) // Nature reviews. Molecular cell biology. — 2007. — Vol. 8, no. 3. — P. 185—194. — doi:10.1038/nrm2122. — PMID 17318224.
Литература
[править | править код]- David L. Nelson, Michael M. Cox. Lehninger Principles of biochemistry. — Fifth edition. — New York: W. H. Freeman and company, 2008. — P. 386. — 1158 p. — ISBN 978-5-4344-0113-5.
- Б. Альбертс, А. Джонсон, Д. Льюис и др. Молекулярная биология клетки / Перевод с английского - А.Н. Дьяконовой, А.В. Дюбы и А.А. Светлова. Под ред. - Е.С. Шилова, Б.П. Копнина, М.А. Лагарьковой, Д.В. Купраша.. — М.—Ижевск: НИЦ «Регулярная и хаотическая динамика», 2013. — Т. 2. — С. 1213. — 1052 с. — ISBN 978-5-4344-0137-1.
- Thomas C. M., Smart E. J. Caveolae structure and function. (англ.) // Journal of cellular and molecular medicine. — 2008. — Vol. 12, no. 3. — P. 796—809. — doi:10.1111/j.1582-4934.2008.00295.x. — PMID 18315571.
Структуры клеточной мембраны | |
|---|---|
| Мембранные липиды[англ.] | |
| Мембранные белки | |
| Другое | |
Эта статья входит в число добротных статей русскоязычного раздела Википедии. |