
 French
French Deutsch
DeutschAbiyogenez - Vikipedi

Biyolojide abiyogenez (a- 'değil' + Yunanca bios 'yaşam' + genesis 'köken') veya yaşamın kökeni, yaşamın basit organik bileşikler gibi cansız maddelerden ortaya çıktığı doğal süreçtir. Hakim bilimsel hipotez, Dünya'da cansız varlıklardan canlı varlıklara geçişin tek bir olay değil, yaşanabilir bir gezegenin oluşumu, organik moleküllerin prebiyotik sentezi, moleküler kendini kopyalama, kendini birleştirme, otokataliz ve hücre zarlarının ortaya çıkışını içeren artan karmaşıklıkta bir süreç olduğudur. Sürecin farklı aşamaları için birçok öneri yapılmıştır.
Abiyogenez çalışması, yaşam öncesi kimyasal reaksiyonların bugün Dünya'dakinden çarpıcı biçimde farklı koşullar altında yaşamı nasıl ortaya çıkardığını belirlemeyi amaçlamaktadır. Öncelikle biyoloji ve kimyanın araçlarını kullanır, daha yeni yaklaşımlar ise birçok bilimin sentezini yapmaya çalışır. Yaşam, karbon ve suyun özelleşmiş kimyası aracılığıyla işler ve büyük ölçüde dört temel kimyasal ailesine dayanır: hücre zarları için lipitler, şekerler gibi karbonhidratlar, protein metabolizması için amino asitler ve kalıtım mekanizmaları için nükleik asit DNA ve RNA. Başarılı bir abiyogenez teorisi, bu molekül sınıflarının kökenlerini ve etkileşimlerini açıklamalıdır. Abiyogeneze yönelik pek çok yaklaşım, kendini kopyalayan moleküllerin ya da bileşenlerinin nasıl ortaya çıktığını araştırmaktadır. Araştırmacılar genellikle mevcut yaşamın bir RNA dünyasından türediğini düşünmektedir, ancak diğer kendi kendini kopyalayan moleküller RNA'dan önce var olmuş olabilir.
Klasik 1952 Miller-Urey deneyi, proteinlerin kimyasal bileşenleri olan amino asitlerin çoğunun, Dünya'nın ilk zamanlarını taklit etmeye yönelik koşullar altında inorganik bileşiklerden sentezlenebileceğini göstermiştir. Yıldırım, radyasyon, mikro meteorların atmosferik girişleri ve deniz ve okyanus dalgalarındaki kabarcıkların patlaması gibi dış enerji kaynakları bu reaksiyonları tetiklemiş olabilir. Diğer yaklaşımlar ("önce metabolizma" hipotezleri) Dünya'nın ilk zamanlarındaki kimyasal sistemlerdeki katalizin kendi kendini kopyalamak için gerekli öncü molekülleri nasıl sağlamış olabileceğini anlamaya odaklanmaktadır.
Genomik bir yaklaşım, yaşamın iki ana dalının üyeleri olan arkea ve bakteriler tarafından paylaşılan genleri tanımlayarak modern organizmaların son evrensel ortak atasını (LUCA) karakterize etmeye çalışmıştır (burada ökaryotlar iki üst âlemli sistemdeki arkea dalına aittir). 355 genin tüm yaşam için ortak olduğu görülmektedir; bu genlerin doğası, LUCA'nın Wood-Ljungdahl yolu ile anaerobik olduğunu, kemiosmoz ile enerji elde ettiğini ve DNA, genetik kod ve ribozomlar ile kalıtsal materyalini koruduğunu ima etmektedir. LUCA 4 milyar yıl (4 Gya) önce yaşamış olmasına rağmen, araştırmacılar onun yaşamın ilk formu olduğuna inanmamaktadırlar. Daha önceki hücreler sızdıran bir zara sahip olabilir ve derin denizlerdeki beyaz dumanlı bir hidrotermal bacanın yakınında doğal olarak oluşan bir proton gradyanından güç almış olabilir.
Dünya, evrende yaşam barındırdığı bilinen tek yer olmaya devam etmektedir ve Dünya'dan elde edilen fosil kanıtlar, abiyogenez çalışmalarının çoğunu bilgilendirmektedir. Dünya 4,54 Gya'da oluşmuştur; Dünya'daki yaşamın tartışmasız en eski kanıtı en az 3,5 Gya'dan kalmadır. Fosil mikroorganizmaların, Hadeen sırasında 4.4 Gya okyanus oluşumundan kısa bir süre sonra, Quebec'ten 3.77 ile 4.28 Gya'ya tarihlenen hidrotermal havalandırma çökeltileri içinde yaşadığı görülmektedir.
Genel bakış
[değiştir | kaynağı değiştir]
Yaşam (kalıtsal) varyasyonlarla üremeden oluşur.[3] NASA yaşamı "Darwinci [yani biyolojik] evrim geçirebilen, kendi kendini idame ettiren kimyasal bir sistem" olarak tanımlamaktadır.[4] Böyle bir sistem karmaşıktır; muhtemelen yaklaşık 4 milyar yıl önce yaşamış tek hücreli bir organizma olan son evrensel ortak ata (LUCA), bugün evrensel olan DNA genetik kodunda kodlanmış yüzlerce gene zaten sahipti. Bu da mesajcı RNA, taşıyıcı RNA ve kodu proteinlere çevirmek için ribozomları içeren bir dizi hücresel mekanizma anlamına gelmektedir. Bu proteinler, Wood-Ljungdahl metabolik yolu aracılığıyla anaerobik solunumunu çalıştıracak enzimleri ve genetik materyalini çoğaltacak bir DNA polimerazı içeriyordu.[5][6]
Abiyogenez (yaşamın kökeni)[7][8][9] araştırmacılarının önündeki zorluk, ilk bakışta tüm parçaları işlevini yerine getirebilmesi için gerekli olan böylesine karmaşık ve birbirine sıkı sıkıya bağlı bir sistemin evrimsel adımlarla nasıl gelişebildiğini açıklamaktır. Örneğin, ister LUCA ister modern bir organizma olsun, bir hücre DNA'sını DNA polimeraz enzimi ile kopyalar ve bu enzim de DNA'daki DNA polimeraz geninin transle edilmesiyle üretilir. Biri olmadan ne enzim ne de DNA üretilebilir.[10] Evrimsel süreç, moleküler kendi kendini kopyalama, hücre zarları gibi kendini birleştirme ve otokatalizi içermiş olabilir.[5][6][11] Bununla birlikte, yaşam olmayanın yaşama geçişi hiçbir zaman deneysel olarak gözlemlenmemiştir.[12]
LUCA gibi canlı bir hücrenin gelişiminin öncülleri, ayrıntıları tartışmalı olsa da yeterince açıktır: mineral ve sıvı su kaynağı ile yaşanabilir bir dünya oluşur. Prebiyotik sentez, proteinler ve RNA gibi polimerler halinde bir araya getirilen bir dizi basit organik bileşik yaratır. LUCA'dan sonraki süreç de kolayca anlaşılabilir: biyolojik evrim, çeşitli formlara ve biyokimyasal yeteneklere sahip çok çeşitli türlerin gelişmesine neden olmuştur. Bununla birlikte, LUCA gibi canlıların basit bileşenlerden türetilmesi anlaşılmaktan uzaktır.[1]
Dünya yaşamın bilindiği tek yer olmaya devam etse de[13][14] astrobiyoloji bilimi diğer gezegenlerde yaşam olduğuna dair kanıtlar aramaktadır. NASA'nın yaşamın kökenine ilişkin 2015 stratejisi, evrimleşebilen makromoleküler sistemlerin çeşitliliğine, seçilimine ve çoğalmasına katkıda bulunan etkileşimleri, aracı yapıları ve işlevleri, enerji kaynaklarını ve çevresel faktörleri belirleyerek ve potansiyel ilkel bilgi polimerlerinin kimyasal manzarasını haritalandırarak bulmacayı çözmeyi amaçlamıştır.[2] Çoğalabilen, genetik bilgi depolayabilen ve seçilime tabi özellikler sergileyebilen polimerlerin ortaya çıkışının, büyük olasılıkla prebiyotik kimyasal evrimin ortaya çıkışında kritik bir adım olduğu öne sürülmüştür.[2] Bu polimerler de çevredeki reaksiyonlarla oluşmuş olabilecek nükleobazlar, amino asitler ve şekerler gibi basit organik bileşiklerden türemiştir.[8][15][16][17] Yaşamın kökenine ilişkin başarılı bir teori, tüm bu kimyasalların nasıl ortaya çıktığını açıklamalıdır.[18]
1960'lara kadar kavramsal tarihçe
[değiştir | kaynağı değiştir]
Kendiliğinden oluşum
[değiştir | kaynağı değiştir]Aristoteles'ten 19. yüzyıla kadar yaşamın kökenine ilişkin eski görüşlerden biri kendiliğinden oluşumdur.[19] Bu teori, "aşağı" hayvanların çürüyen organik maddeler tarafından üretildiğini ve yaşamın tesadüfen ortaya çıktığını savunuyordu.[20][21] Bu teori 17. yüzyıldan itibaren Thomas Browne'un Pseudodoxia Epidemica'sı gibi eserlerde sorgulanmıştır.[22][23] 1665 yılında Robert Hooke bir mikroorganizmanın ilk çizimlerini yayınladı. 1676'da Antonie van Leeuwenhoek, muhtemelen protozoa ve bakteri olan mikroorganizmaları çizdi ve tanımladı.[24] Van Leeuwenhoek kendiliğinden oluşuma karşı çıktı ve 1680'lerde kapalı ve açık et inkübasyonundan böcek üremesinin yakından incelenmesine kadar çeşitli deneyler kullanarak teorinin yanlış olduğuna kendini ikna etti.[25] 1668 yılında Francesco Redi, sineklerin yumurtlaması engellendiğinde ette kurtçuk oluşmadığını göstermiştir.[26] 19. yüzyılın ortalarına gelindiğinde, kendiliğinden oluşumun kanıtlanmadığı düşünülüyordu.[27][28]
Panspermia
[değiştir | kaynağı değiştir]MÖ 5. yüzyılda Anaksagoras'a kadar uzanan bir başka eski fikir de panspermia,[29] yani yaşamın meteoroitler, asteroitler, kuyruklu yıldızlar[30] ve gezegenimsiler[31] tarafından dağıtılmış olarak evrenin her yerinde var olduğu fikridir. Yaşamın kendi içinde nasıl ortaya çıktığını açıklamaya çalışmaz, ancak Dünya'daki yaşamın kökenini başka bir gök cismine kaydırır. Bunun avantajı, yaşamın oluştuğu her bir gezegende değil, daha sınırlı bir dizi yerde (hatta potansiyel olarak tek bir yerde) oluşmuş olması ve daha sonra kuyruklu yıldız veya meteor çarpması yoluyla galaksideki diğer yıldız sistemlerine yayılmış olmasıdır.[32]
"Sıcak küçük bir gölet": ilkel çorba
[değiştir | kaynağı değiştir]Yaşamın cansız maddelerden yavaş aşamalarla oluştuğu fikri, Herbert Spencer'ın 1864-1867 yılları arasında yayımlanan "Biyolojinin İlkeleri" adlı kitabında ve William Turner Thiselton-Dyer'ın 1879 tarihli "Kendiliğinden Oluşum ve Evrim Üzerine" adlı makalesinde yer almıştır. Charles Darwin 1 Şubat 1871'de Joseph Hooker'a bu yayınlar hakkında yazdı ve kendi spekülasyonunu ortaya koyarak yaşamın ilk kıvılcımının "her türlü amonyak ve fosforik tuzun, ışığın, ısının, elektriğin vb. bulunduğu, daha karmaşık değişimlere uğramaya hazır bir protein bileşiğinin kimyasal olarak oluştuğu sıcak küçük bir havuzda" başlamış olabileceğini öne sürdü. Darwin, "günümüzde böyle bir maddenin anında yutulacağını ya da emileceğini, oysa canlılar oluşmadan önce böyle bir şeyin söz konusu olamayacağını" açıklamaya devam etmiştir.[33][34][35]
1924'te Alesandr Oparin ve 1929'da J. B. S. Haldane, ilk hücreleri oluşturan ilk moleküllerin ilkel bir çorbadan yavaşça kendi kendine organize olduğunu öne sürmüş ve bu teori Oparin-Haldane hipotezi olarak adlandırılmıştır.[36][37] Haldane, Dünya'nın prebiyotik okyanuslarının organik bileşiklerin oluşabileceği "sıcak seyreltik bir çorbadan" oluştuğunu öne sürmüştür.[21][38] J. D. Bernal, bu tür mekanizmaların yaşam için gerekli moleküllerin çoğunu inorganik öncülerden oluşturabileceğini gösterdi.[39] 1967'de üç "aşama" önerdi: biyolojik monomerlerin kökeni; biyolojik polimerlerin kökeni; ve moleküllerden hücrelere evrim.[40][41]
Miller-Urey deneyi
[değiştir | kaynağı değiştir]1952 yılında Stanley Miller ve Harold Urey, Oparin-Haldane hipotezinin öne sürdüğü gibi prebiyotik koşullar altında organik moleküllerin inorganik öncüllerden kendiliğinden nasıl oluşabileceğini göstermek için kimyasal bir deney gerçekleştirmiştir. Amino asitler gibi basit organik monomerleri oluşturmak için metan, amonyak ve hidrojenin yanı sıra su buharı gibi yüksek oranda indirgeyici (oksijenden yoksun) bir gaz karışımı kullandı.[42][43] Bernal, Miller-Urey deneyi için "bu tür moleküllerin oluşumunu açıklamak yeterli değildir, gerekli olan, bu moleküllerin kökenlerinin, serbest enerji için uygun kaynakların ve yutakların varlığını öne süren fiziksel-kimyasal bir açıklamasıdır" demiştir.[44] Bununla birlikte, mevcut bilimsel fikir birliği ilkel atmosferi zayıf indirgeyici veya nötr olarak tanımlamakta, bu da üretilebilecek amino asitlerin miktarını ve çeşitliliğini azaltmaktadır.[45][46] Bununla birlikte, erken okyanuslarda bulunan demir ve karbonat minerallerinin eklenmesi, çok çeşitli amino asitler üretmektedir.[45] Daha sonraki çalışmalar diğer iki potansiyel indirgeyici ortama odaklanmıştır: dış uzay ve derin deniz hidrotermal bacaları.[47][48][49]
Yaşanabilir bir Dünya üretmek
[değiştir | kaynağı değiştir]Yaşamın kökeni zaman çizelgesi | ||||||||||||||||||||||||
−13 — – −12 — – −11 — – −10 — – −9 — – −8 — – −7 — – −6 — – −5 — – −4 — – −3 — – −2 — – −1 — – 0 — |
| |||||||||||||||||||||||
İlk yıldızlarla birlikte erken evren
[değiştir | kaynağı değiştir]Yaklaşık 14 Gya'da meydana gelen Büyük Patlama'dan kısa bir süre sonra, evrende mevcut olan tek kimyasal elementler periyodik tablodaki en hafif üç atom olan hidrojen, helyum ve lityumdu. Bu elementler yavaş yavaş bir araya gelerek yıldızları oluşturdu. Bu ilk yıldızlar büyük kütleli ve kısa ömürlüydü ve yıldız nükleosentezi yoluyla tüm ağır elementleri üretiyorlardı. Şu anda evrende en bol bulunan dördüncü kimyasal element olan karbon (hidrojen, helyum ve oksijenden sonra), esas olarak beyaz cüce yıldızlarda, özellikle de Güneş'in kütlesinin iki katından daha büyük olanlarda oluşmuştur.[50]
İlk yıldızlar büyük kütleli ve kısa ömürlüydü ve yıldız nükleosentezi yoluyla tüm ağır elementleri üretirlerdi. Yıldız nükleosentezi yoluyla element oluşumu en kararlı element olan Demir-56'ya kadar ilerler. Daha ağır elementler, bir yıldızın yaşam döngüsünün sonundaki süpernovalar sırasında oluşmuştur. Şu anda evrende en bol bulunan dördüncü kimyasal element olan karbon (hidrojen, helyum ve oksijenden sonra), esas olarak beyaz cüce yıldızlarda, özellikle de Güneş'in kütlesinin iki katından daha büyük olanlarda oluşmuştur.[51] Bu yıldızlar yaşam döngülerinin sonuna ulaştıklarında, aralarında karbon ve oksijenin de bulunduğu bu daha ağır elementleri evrenin dört bir yanına fırlattılar. Bu daha ağır elementler, kayalık gezegenler ve diğer cisimler de dahil olmak üzere yeni cisimlerin oluşumuna olanak sağladı.[52] Bulutsu hipotezine göre, Güneş Sistemi'nin oluşumu ve evrimi 4,6 Gya'da dev bir moleküler bulutun küçük bir bölümünün kütleçekimsel çöküşüyle başladı. Çöken kütlenin çoğu merkezde toplanarak Güneş'i oluştururken, geri kalanı düzleşerek gezegenlerin, uyduların, asteroitlerin ve diğer küçük Güneş Sistemi cisimlerinin oluştuğu bir ön gezegen diskine dönüştü.[53]
Dünya'nın ortaya çıkışı
[değiştir | kaynağı değiştir]Dünya 4.54 Gya'da oluşmuştur.[54][55] Hadeen Dünyası (oluşumundan 4 Gya'ya kadar) ilk başta herhangi bir canlı organizma için uygun değildi. Oluşumu sırasında Dünya başlangıçtaki kütlesinin önemli bir kısmını kaybetti ve sonuç olarak moleküler hidrojeni ve orijinal inert gazların büyük kısmını tutacak kütleçekiminden yoksun kaldı.[56] Dünya'nın 4.48 Ga'da ilk birikiminden kısa bir süre sonra, varsayılan bir çarpıştırıcı olan Theia ile çarpışmasının, sonunda Ay'ı oluşturacak olan fırlatılmış enkazı yarattığı düşünülmektedir.[57] Bu çarpışma Dünya'nın birincil atmosferini ortadan kaldırmış ve geride viskoz silikat ve karbondioksit bulutları bırakmıştır. Bu kararsız atmosfer kısa ömürlü olmuş ve kısa bir süre sonra yoğunlaşarak silikat Dünya'yı oluşturmuş, geride büyük ölçüde su buharı, nitrojen ve karbondioksitten oluşan, daha az miktarda karbon monoksit, hidrojen ve kükürt bileşikleri içeren bir atmosfer bırakmıştır.[58][59] Karbondioksitin sudaki çözeltisinin, denizleri yaklaşık 5,5 pH ile hafif asidik hale getirdiği düşünülmektedir.[60]
Okyanuslar, Dünya'nın oluşumundan 200 milyon yıl kadar sonra, kaynama noktasına yakın (100 C) indirgeyici bir ortamda, 5.8'lik pH hızla nötre doğru yükselirken ortaya çıkmış olabilir.[61] Bu senaryo, Batı Avustralya'daki Narryer Dağı'nın metamorfize kuvarsitinden elde edilen 4.404 Gya zirkon kristallerinin tarihlendirilmesinden destek bulmuştur.[62] Muhtemelen artan volkanizmaya rağmen, Dünya 4.4 ila 4.3 Gya arasında, kıtasal kabuğun çok az olduğu, çalkantılı bir atmosfere ve T Tauri evresindeki Güneş'ten, kozmik radyasyondan ve devam eden asteroit ve kuyruklu yıldız çarpmalarından kaynaklanan yoğun ultraviyole ışığa maruz kalan bir hidrosfere sahip bir su dünyası olabilir.[63]
Geç Dönem Ağır Bombardıman hipotezi, 4,28[64] ve 3,8 Gya arasındaki Hadeen ortamının yaşam için oldukça tehlikeli olduğunu öne sürmektedir. Nice modelini takiben, dev gezegenlerin yörüngelerindeki değişiklikler Dünya'yı, Ay'ı ve iç gezegenleri çukurlaştıran asteroit ve kuyruklu yıldızlarla bombardımana tutmuş olabilir.[65] Sık çarpışmalar fotosentezi gerçekleşmez hale getirebilirdi.[59][66][67][68] Bu tür yıkıcı olaylar arasındaki dönemler, erken ortamlarda yaşamın olası kökeni için zaman pencereleri verir. Eğer yaşamın ortaya çıktığı yer derin deniz hidrotermal ortamı ise abiyogenez 4.0-4.2 Gya kadar erken bir zamanda gerçekleşmiş olabilir. Eğer bölge Dünya yüzeyindeyse abiyogenez sadece 3.7 ile 4.0 Gya arasında gerçekleşmiş olabilir.[69] Ancak, yeni Ay araştırmaları ve örnekleri, Nice modelinin mimarlarından biri de dahil olmak üzere bilim insanlarının Geç Dönem Ağır Bombardıman'ın önemini vurgulamalarına yol açmıştır.[70]
Eğer yaşam okyanusta on metreden daha derinlerde evrimleştiyse hem geç darbelerden hem de o zamanlar güneşten gelen yüksek seviyelerdeki ultraviyole radyasyondan korunmuş olacaktır. Jeotermal olarak ısıtılan okyanus kabuğu, derin hidrotermal bacalar yoluyla Miller-Urey deneylerinin gösterdiğinden çok daha fazla organik bileşik üretmiş olabilir.[71] Mevcut enerji, hipertermofilik bakterilerin ve termoasidofilik arkeaların yaşadığı sıcaklıklar olan 100-150 °C'de en üst düzeye çıkar. Bu modern organizmalar, LUCA'nın hayatta kalan en yakın akrabaları arasında olabilir.[72]
Yaşamın en eski kanıtı
[değiştir | kaynağı değiştir]Yaşam, erimiş Hadeen'in ardından yeterli kabuğun katılaştığı Eoarkeen döneminde, 3,5 Gya'dan daha uzun bir süre[73][74][75] Dünya'da var olmuştur.[76][77][78] Şimdiye kadar bulunan en eski fiziksel yaşam kanıtı, Kuzey Quebec'in Nuvvuagittuq Yeşiltaş Kuşağı'nda, en az 3.77 ve muhtemelen 4.28 Gya'da bantlı demir formasyonu kayalarındaki mikrofosillerden oluşmaktadır. Mikroorganizmalar, Hadeen sırasında okyanusların 4.4 Gya oluşumundan kısa bir süre sonra hidrotermal menfez çökeltileri içinde yaşamıştır. Mikroplar modern hidrotermal havalandırma bakterilerine benzemektedir ve abiyogenezin böyle bir ortamda başladığı görüşünü desteklemektedir.[64]
Biyojenik grafit, güneybatı Grönland'dan[79] 3.7 Gya metasedimenter kayalarda ve 3.49 Gya Batı Avustralya kumtaşından mikrobiyal mat fosillerinde bulunmuştur.[80] Güneybatı Grönland'daki Isua suprakrustal kuşağı yakınlarındaki Akilia Adası'ndan 3.7 Gya'ya tarihlenen kayalarda erken yaşamın kanıtları biyojenik karbon izotopları göstermiştir.[81] Isua suprakrustal kuşağının diğer kısımlarında, garnet kristalleri içinde hapsolmuş grafit kalıntıları yaşamın diğer elementleriyle bağlantılıdır: oksijen, azot ve muhtemelen fosfat formundaki fosfor, 3.7 Gya yaşamı için daha fazla kanıt sağlar.[82] Batı Avustralya'nın Pilbara bölgesinde, fosilleşmiş bir plajdaki pirit içeren kumtaşında, oksijen yokluğunda fotosentez yoluyla sülfürü oksitleyen yuvarlak boru şeklindeki hücrelerle erken yaşamın ikna edici kanıtları bulunmuştur.[83][84] Batı Avustralya'dan elde edilen zirkonlar, Dünya'da yaşamın en az 4.1 Gya'da var olduğunu göstermektedir.[85]
Batı Avustralya'nın Pilbara bölgesi, stromatolit adı verilen katmanlı yapılar da dahil olmak üzere 3.48 Gya kayaları içeren Dresser Formasyonunu içermektedir. Bunların modern benzerleri siyanobakteriler de dahil olmak üzere fotosentetik mikroorganizmalar tarafından oluşturulmuştur.[86] Bunlar deforme olmamış hidrotermal-sedimanter tabakalar içinde yer alır; dokuları biyojenik bir kökene işaret eder. Dresser formasyonunun bazı kısımları karadaki sıcak su kaynaklarını korumaktadır, ancak diğer bölgeler sığ denizler gibi görünmektedir.[87]
-
 Glacier Ulusal Parkı, Siyeh Formasyonu'ndaki stromatolitler 3,5 Gya'ya tarihlendirilerek en eski yaşam formları arasına yerleştirilmiştir
Glacier Ulusal Parkı, Siyeh Formasyonu'ndaki stromatolitler 3,5 Gya'ya tarihlendirilerek en eski yaşam formları arasına yerleştirilmiştir -
 Shark Bay'deki modern stromatolitler, fotosentetik siyanobakteriler tarafından oluşturulmuştur
Shark Bay'deki modern stromatolitler, fotosentetik siyanobakteriler tarafından oluşturulmuştur


Moleküllerin üretilmesi: prebiyotik sentez
[değiştir | kaynağı değiştir]Hidrojen ve helyum dışındaki tüm kimyasal elementler yıldız nükleosentezinden türemiştir. Yaşamın temel kimyasal bileşenleri - karbon-hidrojen molekülü (CH), karbon-hidrojen pozitif iyonu (CH+) ve karbon iyonu (C+) - yıldızlardan gelen ultraviyole ışıkla üretilmiştir.[88] Organik moleküller de dahil olmak üzere karmaşık moleküller hem uzayda hem de gezegenlerde doğal olarak oluşur.[89] Dünya'nın ilk zamanlarındaki organik moleküller ya çarpma şokları ya da ultraviyole ışık, redoks eşleşmesi veya elektrik deşarjları gibi diğer enerji kaynakları tarafından yönlendirilen organik molekül sentezi ile karasal kökenli olabilir; ya da gezegene yağan yıldızlararası toz bulutlarında oluşan organik moleküller ile dünya dışı kökenli (psödo-panspermia) olabilir.[90][91]
Gözlemlenen dünya dışı organik moleküller
[değiştir | kaynağı değiştir]Organik bir bileşik, molekülleri karbon içeren bir kimyasaldır. Karbon Güneş'te, yıldızlarda, kuyruklu yıldızlarda ve çoğu gezegenin atmosferinde bol miktarda bulunur.[92] Organik bileşikler uzayda nispeten yaygındır, moleküler bulutlarda ve çöküntü örtülerinde meydana gelen "karmaşık moleküler sentez fabrikaları" tarafından oluşturulur ve reaksiyonlar çoğunlukla iyonlaştırıcı radyasyon tarafından başlatıldıktan sonra kimyasal olarak gelişir.[89][93][94] Meteorlarda guanin, adenin, sitozin, urasil ve timin gibi pürin ve pirimidin nükleobazları bulunmuştur. Bunlar Dünya'nın ilk zamanlarında DNA ve RNA'nın oluşması için gerekli materyalleri sağlamış olabilir.[95] Amino asit glisin Wild 2 kuyruklu yıldızından fırlatılan materyalde bulunmuştur; daha önce meteorlarda tespit edilmişti.[96] Kuyruklu yıldızlar, iyonlaştırıcı radyasyon altında basit karbon bileşiklerinden oluşan katran benzeri organik bir madde olduğu düşünülen koyu renkli malzemeyle kaplıdır. Kuyruklu yıldızlardan gelen bir malzeme yağmuru bu tür karmaşık organik molekülleri Dünya'ya getirmiş olabilir.[59][97][98] Geç Ağır Bombardıman sırasında meteorların Dünya'ya yılda beş milyon tona kadar organik prebiyotik element getirmiş olabileceği tahmin edilmektedir.[59]
PAH dünya hipotezi
[değiştir | kaynağı değiştir]
Polisiklik aromatik hidrokarbonlar (PAH) gözlemlenebilir evrende en yaygın ve bol bulunan çok atomlu moleküllerdir ve önemli bir karbon deposudur.[92][99][100][101] Büyük Patlama'dan kısa bir süre sonra oluşmuş gibi görünmektedirler[100][101][102] ve yeni yıldızlar ve ötegezegenlerle ilişkilidirler.[92] Dünya'nın ilkel denizinin muhtemel bir bileşenidirler.[100][101][102] PAH'lar bulutsularda,[103] yıldızlararası ortamda, kuyruklu yıldızlarda ve meteoritlerde tespit edilmiştir.[92]
PAH dünyası hipotezi, PAH'ları RNA dünyasının öncülleri olarak ortaya koymaktadır.[104] Yaşamının erken dönemlerinde Güneş'e benzeyen HH 46-IR adlı bir yıldız, siyanür bileşikleri, hidrokarbonlar ve karbonmonoksit gibi moleküller içeren bir malzeme diski ile çevrilidir. Yıldızlararası ortamdaki PAH'lar hidrojenasyon, oksijenasyon ve hidroksilasyon yoluyla canlı hücrelerde kullanılan daha karmaşık organik bileşiklere dönüştürülebilir.[105]
Nükleobazlar
[değiştir | kaynağı değiştir]Yıldızlararası toz partikülleri tarafından Dünya'ya getirilen organik bileşiklerin çoğu, kendilerine özgü yüzey-katalitik aktiviteleri sayesinde karmaşık moleküllerin oluşmasına yardımcı olmuştur.[106][107] Murchison meteoritindeki organik bileşiklerin 12C/13C izotopik oranları üzerine yapılan çalışmalar, RNA bileşeni urasil ve ksantin de dahil olmak üzere ilgili moleküllerin dünya dışında oluştuğunu göstermektedir.[108] NASA'nın meteoritler üzerinde yaptığı çalışmalar, dört DNA nükleobazının (adenin, guanin ve ilgili organik moleküller) da uzayda oluştuğunu göstermektedir.[106][109][110] Evrene nüfuz eden kozmik toz, yıldızlar tarafından hızla yaratılabilecek karmaşık organikler ("karışık aromatik-alifatik yapıya sahip amorf organik katılar") içerir.[111] Bir şeker molekülü ve RNA öncüsü olan glikolaldehit, protostarların çevresi ve meteoritler de dahil olmak üzere uzay bölgelerinde tespit edilmiştir.[112][113]
Laboratuvar sentezi
[değiştir | kaynağı değiştir]1860'ların başlarında yapılan deneyler, basit karbon kaynaklarının bol miktarda inorganik katalizörle etkileşiminden biyolojik olarak ilgili moleküllerin üretilebileceğini göstermiştir. "Çorba" teorisinin öne sürdüğü koşullar altında abiyotik olarak üretilen monomerlerden karmaşık polimerlerin kendiliğinden oluşması kolay değildir. Gerekli temel organik monomerlerin yanı sıra, Miller-Urey ve Joan Oró deneyleri sırasında polimer oluşumunu engelleyecek bileşikler de yüksek konsantrasyonlarda oluşmuştur.[114] Biyoloji, kodlanmış protein enzimleri için esasen 20 amino asit kullanır ve bu da yapısal olarak mümkün olan ürünlerin çok küçük bir alt kümesini temsil eder. Yaşam mevcut olanı kullanma eğiliminde olduğundan, kullanılan kümenin neden bu kadar küçük olduğuna dair bir açıklamaya ihtiyaç vardır.[115]
Şekerler
[değiştir | kaynağı değiştir]
Alexander Butlerov 1861'de formaldehitin bazik koşullar altında kalsiyum gibi divalent metal iyonları ile ısıtıldığında formoz reaksiyonunun tetrozlar, pentozlar ve heksozlar dahil şekerler oluşturduğunu göstermiştir. R. Breslow 1959'da reaksiyonun otokatalitik olduğunu öne sürmüştür.[116]
Nükleobazlar
[değiştir | kaynağı değiştir]Guanin ve adenin gibi nükleobazlar, hidrojen siyanür (HCN) ve amonyak gibi basit karbon ve nitrojen kaynaklarından sentezlenebilir.[117] Formamid, karasal minerallerle ısıtıldığında dört ribonükleotidin tamamını üretir. Formamid evrende her yerde bulunur ve su ile HCN'nin reaksiyonuyla üretilir. Suyun buharlaştırılmasıyla konsantre edilebilir.[118][119] HCN sadece henüz var olmayan aerobik organizmalar (ökaryotlar ve aerobik bakteriler) için zehirlidir. Glisin amino asidinin sentezi gibi diğer kimyasal süreçlerde rol oynayabilir.[59]
Urasil, sitozin ve timin gibi DNA ve RNA bileşenleri, meteoritlerde bulunan pirimidin gibi başlangıç kimyasalları kullanılarak uzay koşullarında sentezlenebilir. Pirimidin kırmızı dev yıldızlarda ya da yıldızlararası toz ve gaz bulutlarında oluşmuş olabilir.[120] Dört RNA bazının tümü, dünya dışı çarpmalar gibi yüksek enerji yoğunluklu olaylarda formamitten sentezlenebilir.[121]
İnorganik malzemelerden baz sentezlemek için başka yollar da rapor edilmiştir.[122] Donma sıcaklıkları, hidrojen siyanür gibi temel öncüller için yoğunlaştırma etkisi nedeniyle pürinlerin sentezi için avantajlıdır.[123] Bununla birlikte, adenin ve guanin sentez için donma koşulları gerektirirken, sitozin ve urasil kaynama sıcaklıkları gerektirebilir.[124] Amonyak ve siyanür 25 yıl boyunca bir dondurucuda bırakıldığında buzda yedi amino asit ve on bir tür nükleobaz oluşmuştur.[125][126] S-triazinler (alternatif nükleobazlar), pirimidinler ve adenin, bir üre çözeltisi indirgeyici bir atmosfer altında donma-çözülme döngülerine tabi tutularak ve enerji kaynağı olarak kıvılcım deşarjları kullanılarak sentezlenebilir.[127] Bu kadar düşük bir sıcaklıkta bu reaksiyonların olağandışı hızına getirilen açıklama, buz içindeki mikroskobik sıvı ceplerinde safsızlıkları toplayarak moleküllerin daha sık çarpışmasına neden olan ötektik dondurmadır.[128]
Uygun veziküllerin üretilmesi
[değiştir | kaynağı değiştir]
Lipit dünyası teorisi, kendi kendini kopyalayan ilk nesnenin lipit benzeri olduğunu varsayar.[129][130] Fosfolipitler suda çalkalanırken lipit çift tabakaları oluştururlar - hücre zarlarında olduğu gibi aynı yapıdır. Bu moleküller Dünya'nın erken dönemlerinde mevcut değildi, ancak diğer amfifilik uzun zincirli moleküller de zarlar oluşturur. Bu cisimler ilave lipitlerin eklenmesiyle genişleyebilir ve kendiliğinden benzer boyut ve bileşimde iki yavruya bölünebilir. Ana fikir, lipit gövdelerinin moleküler bileşiminin bilgi depolamanın bir ön hazırlığı olduğu ve evrimin bilgi depolayan RNA gibi polimerlerin ortaya çıkmasına yol açtığıdır. Prebiyotik dünyada var olmuş olabilecek amfifillerden veziküller üzerine yapılan çalışmalar şimdiye kadar bir ya da iki tip amfifilden oluşan sistemlerle sınırlı kalmıştır.[131]
Bir lipit çift katmanlı zarı, amfifillerin çok sayıda kombinasyonundan oluşabilir. Bunların en iyisi bir hiper döngünün, aslında bir zar bölgesi ve vezikül içinde hapsolmuş belirli bir bileşik tarafından temsil edilen iki karşılıklı katalizörden oluşan bir pozitif geri beslemenin oluşumunu tercih ederdi.[132][133] Bu tür bölge/bileşik çiftleri yavru veziküllere aktarılabilir ve bu da doğal seçilime izin verecek farklı vezikül soylarının ortaya çıkmasına neden olur.[134]
Bir protosel, yaşamın kökenine bir basamak taşı olarak önerilen, kendi kendine organize olmuş, küresel bir lipit topluluğudur.[131] Klasik tersinmez termodinamik teorisi, genelleştirilmiş bir kimyasal potansiyel altında kendi kendine birleşmeyi dağıtıcı sistemler çerçevesinde ele alır.[135][136][137]
Evrimde temel bir soru, basit protosellerin ilk olarak nasıl ortaya çıktığı ve bir sonraki nesle üreme katkısı açısından nasıl farklılaştığı ve böylece yaşamın evrimini nasıl yönlendirdiğidir. İşlevsel bir protosel (2014 itibarıyla) henüz laboratuvar ortamında elde edilememiştir.[138][139][140] Kendiliğinden bir araya gelen veziküller ilkel hücrelerin temel bileşenleridir.[131] Termodinamiğin ikinci yasası, evrenin entropinin arttığı bir yönde hareket etmesini gerektirir, ancak yaşam, büyük organizasyon derecesi ile ayırt edilir. Bu nedenle, yaşam süreçlerini cansız maddeden ayırmak için bir sınıra ihtiyaç vardır.[141] Irene Chen ve Jack W. Szostak, temel protosellerin, diferansiyel üreme, rekabet ve enerji depolamanın ilkel biçimleri de dahil olmak üzere hücresel davranışlara yol açabileceğini öne sürmektedir.[139] Membran molekülleri için rekabet, stabilize membranları destekleyecek, çapraz bağlı yağ asitlerinin ve hatta günümüzün fosfolipitlerinin evrimi için seçici bir avantaj sağlayacaktır.[139] Bu tür bir mikrokapsülleme, büyük biyomolekülleri içeride tutarken, zar içinde metabolizmaya ve küçük moleküllerin değişimine izin verecektir. Böyle bir zar, bir hücrenin iyonları zar boyunca pompalayarak enerji depolamak üzere kendi elektrokimyasal gradyanını yaratması için gereklidir.[142][143]
Enerji ve entropi
[değiştir | kaynağı değiştir]Yaşam, moleküller kendilerini canlı madde olarak organize ettiklerinde entropi ya da düzensizlik kaybını gerektirir. Yaşamın ortaya çıkması ve karmaşıklığın artması, genel entropinin asla azalmayacağını belirten termodinamiğin ikinci yasasıyla çelişmez, çünkü canlı bir organizma başka yerlerde entropinin artması pahasına (örneğin ısı ve atık üretimi) bazı yerlerde (örneğin canlı vücudu) düzen yaratır.[144][145][146]
Dünya'nın ilk zamanlarında kimyasal reaksiyonlar için birden fazla enerji kaynağı mevcuttu. Jeotermal süreçlerden gelen ısı, kimya için standart bir enerji kaynağıdır. Diğer örnekler arasında güneş ışığı, şimşek,[59] mikrometeorların[147] atmosferik girişleri ve deniz ve okyanus dalgalarındaki kabarcıkların patlaması sayılabilir.[148] Bu durum deneyler[149][150] ve simülasyonlarla doğrulanmıştır.[151] Demir-kükürt kimyasında olduğu gibi, elverişsiz reaksiyonlar son derece elverişli reaksiyonlar tarafından yönlendirilebilir. Örneğin bu, muhtemelen karbon fiksasyonu için önemliydi.[a] Demir-sülfür kimyası yoluyla CO2'nin H2S ile reaksiyonu yoluyla karbon fiksasyonu elverişlidir ve nötr pH ve 100 °C'de gerçekleşir. Hidrotermal bacaların yakınında bol miktarda bulunan demir-kükürt yüzeyleri, az miktarda amino asit ve diğer biyomoleküllerin üretimini sağlayabilir.[59]
Kemiosmoz
[değiştir | kaynağı değiştir]
1961 yılında Peter Mitchell, bir hücrenin birincil enerji dönüşüm sistemi olarak kemiosmozu önermiştir. Günümüzde canlı hücrelerde her yerde bulunan bu mekanizma, mikroorganizmalarda ve ökaryotların mitokondrilerinde enerji dönüşümünü sağlayarak yaşamın erken dönemleri için olası bir aday haline gelmiştir.[152][153] Mitokondri, kimyasal sentezler gibi hücresel süreçleri yürütmek için kullanılan hücrenin enerji para birimi olan adenozin trifosfat (ATP) üretir. ATP sentez mekanizması, ATP sentaz enziminin gömülü olduğu kapalı bir zarı içerir. Güçlü bir şekilde bağlanmış ATP'yi serbest bırakmak için gereken enerji, membran boyunca hareket eden protonlardan kaynaklanır.[154] Modern hücrelerde bu proton hareketleri, elektrokimyasal bir gradyanı koruyarak iyonların zar boyunca pompalanmasından kaynaklanır. İlk organizmalarda bu gradyan, hidrotermal bir menfezden gelen akış ile çevredeki deniz suyu arasındaki kimyasal bileşim farkından[143] ya da belki de karasal bir kökene sahipse lipid membranları boyunca kemiosmotik enerjinin gelişmesine elverişli meteorik kinonlardan sağlanmış olabilir.[155]

RNA dünyası
[değiştir | kaynağı değiştir]RNA dünyası hipotezi, kendi kendini kopyalayan ve katalitik RNA'ya sahip ancak DNA ya da proteinlerin bulunmadığı erken bir Dünya'yı tanımlamaktadır.[156] Birçok araştırmacı, RNA dünyasının şu anda hakim olan DNA temelli yaşamdan önce gelmiş olması gerektiği konusunda hemfikirdir.[157] Ancak, RNA temelli yaşam ilk var olan yaşam olmayabilir.[158][159] Bir başka model de Darwin'in ıslanma ve kuruma döngülerine sahip "sıcak küçük göletini" yansıtmaktadır.[160]
RNA, translasyon sürecinin merkezinde yer alır. Küçük RNA'lar yaşam için gerekli tüm kimyasal grupları ve bilgi transferlerini katalize edebilir.[159][161] RNA modern organizmalarda genetik bilgiyi hem ifade eder hem de korur; ve RNA'nın kimyasal bileşenleri, bugünkünden çok farklı olan Dünya'nın ilk zamanlarına yakın koşullar altında kolayca sentezlenebilir. Ribozimin yapısı, merkezi bir RNA çekirdeği ve peptit bağı oluşumunu katalize eden aktif bölgenin 18 Å yakınında hiçbir amino asit yan zinciri bulunmaması nedeniyle "dumanı tüten silah" olarak adlandırılmıştır.[158][162][163]
RNA dünyası kavramı 1962 yılında Alexander Rich tarafından ortaya atılmış[164] ve bu terim 1986 yılında Walter Gilbert tarafından kullanılmıştır.[159][165] Sitozin ve urasil nükleotitlerinin abiyotik sentezinin açıklanmasında başlangıçta zorluklar yaşanmıştır.[166] Daha sonraki araştırmalar olası sentez yollarını göstermiştir; örneğin formamid, çeşitli karasal minerallerin varlığında ısıtıldığında dört ribonükleotidin tamamını ve diğer biyolojik molekülleri üretmektedir.[118][119]

RNA replikaz, daha fazla RNA replikasyonu için hem kod hem de katalizör olarak işlev görebilir, yani otokatalitik olabilir. Jack Szostak, bazı katalitik RNA'ların daha küçük RNA dizilerini bir araya getirerek kendi kendini kopyalama potansiyeli yaratabildiğini göstermiştir. Birbirlerinin sentezini katalize eden iki ribozim içeren RNA replikasyon sistemleri, ürünün yaklaşık bir saatlik bir ikiye katlanma süresi göstermiş ve deneysel koşullar altında doğal seçilime tabi olmuştur.[158][167][168] Eğer Dünya'nın erken dönemlerinde bu tür koşullar mevcut olsaydı doğal seçilim bu tür otokatalitik setlerin çoğalmasını destekleyecek ve bunlara başka işlevler de eklenebilecekti.[169][170][171] RNA'nın kendiliğinden bir araya gelmesi hidrotermal bacalarda kendiliğinden gerçekleşebilir.[172][173][174] İlk tRNA formu böyle bir çoğaltıcı molekül halinde bir araya gelmiş olabilir.[175]
Protein sentezinin olası öncülleri arasında kısa peptit kofaktörlerinin sentezi veya RNA'nın kendi kendini katalize eden duplikasyonu yer alır. Her ne kadar bazı roller proteinler tarafından devralınmış olsa da atasal ribozomun tamamen RNA'dan oluşması muhtemeldir. Bu konuda geriye kalan başlıca sorular arasında ribozomun evrimi için seçici gücün tanımlanması ve genetik kodun nasıl ortaya çıktığının belirlenmesi yer almaktadır.[176]
Eugene Koonin, "biyolojik sistemlerin çekirdeğini oluşturan ve biyolojik evrimin görünürdeki ön koşulu olan kilit süreçler olan replikasyon ve translasyonun kökeni için şu anda ikna edici bir senaryo bulunmadığını" savunmuştur. RNA dünyası kavramı bu muammanın çözümü için en iyi şansı sunabilir ancak şu ana kadar etkin bir RNA replikazının veya çeviri sisteminin ortaya çıkışını yeterince açıklayamamaktadır."[177]
Filogeni ve LUCA
[değiştir | kaynağı değiştir]Carl Woese'nin 1977'deki çalışmalarından başlayarak, genomik çalışmalar tüm modern yaşam formlarının son evrensel ortak atasını (LUCA) filogenetik yaşam ağacında bakteriler ile arkealar ve ökaryotlardan oluşan bir kladın arasına yerleştirmiştir. 4 Gya'nın üzerinde yaşamıştır.[178][179] Az sayıda çalışma LUCA'yı bakterilerin içine yerleştirmiş, arkealar ve ökaryotların evrimsel olarak Eubacteria'nın içinden türediğini öne sürmüştür;[180] Thomas Cavalier-Smith fenotipik olarak çeşitlilik gösteren bakteriyel filum Chloroflexota'nın LUCA'yı içerdiğini öne sürmüştür.[181]
-
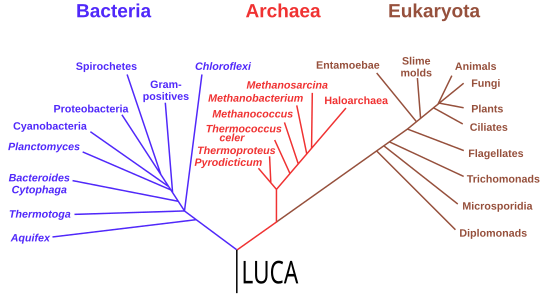 Kökte son evrensel ortak atayı (LUCA) gösteren filogenetik ağaç. Başlıca kladlar bir yanda bakteriler, diğer yanda arkealar ve ökaryotlardır.
Kökte son evrensel ortak atayı (LUCA) gösteren filogenetik ağaç. Başlıca kladlar bir yanda bakteriler, diğer yanda arkealar ve ökaryotlardır.

2016 yılında, LUCA'da bulunması muhtemel 355 genden oluşan bir küme tanımlanmıştır. Bakteri ve arkealarden toplam 6,1 milyon prokaryotik gen dizilenmiş ve LUCA'da muhtemelen ortak olan 286.514 protein kümesi arasından 355 protein kümesi tanımlanmıştır. Sonuçlar, LUCA'nın Wood-Ljungdahl yolağı ile anaerobik, azot ve karbon sabitleyici, termofilik olduğunu göstermektedir. Kofaktörleri hidrojen, karbondioksit, demir ve geçiş metalleri açısından zengin bir ortama bağımlı olduğunu göstermektedir. Genetik materyali muhtemelen DNA idi ve 4 nükleotit genetik kod, mesajcı RNA, taşıyıcı RNA ve kodu enzimler gibi proteinlere çevirmek için ribozomlar gerektiriyordu. LUCA muhtemelen jeokimyasal olarak aktif bir ortamda anaerobik bir hidrotermal baca ortamında yaşıyordu. Belli ki zaten karmaşık bir organizmaydı ve öncülleri olmalıydı; ilk canlı değildi.[10][182] LUCA'nın fizyolojisi tartışmalıdır.[183][184][185]
-
 LUCA sistemleri ve çevresi
LUCA sistemleri ve çevresi

Leslie Orgel, genetik kod için erken çeviri mekanizmasının hata felaketine açık olacağını savunmuştur. Ancak Geoffrey Hoffmann, "Orgel'in paradoksuna" karşı bu tür makinelerin istikrarlı bir şekilde çalışabileceğini göstermiştir.[186][187][188]
Uygun jeolojik ortamlar
[değiştir | kaynağı değiştir]Derin deniz hidrotermal bacaları
[değiştir | kaynağı değiştir]
İlk mikrofosiller, metan, amonyak, karbondioksit ve hidrojen sülfür gibi mevcut yaşam için zehirli olan gazların bulunduğu sıcak bir dünyadan gelmiş olabilir.[189] Yaşam ağacının analizi, termofilik ve hipertermofilik bakteri ve arkeaları köke en yakın yere yerleştirerek yaşamın sıcak bir ortamda evrimleşmiş olabileceğini düşündürmektedir.[190] Derin deniz ya da alkali hidrotermal baca teorisi, yaşamın denizaltı hidrotermal bacalarında başladığını ileri sürmektedir.[191][192] Martin ve Russell, "yaşamın, sülfit bakımından zengin hidrotermal sıvı ile Hadeen okyanus tabanının demir (II) içeren suları arasındaki redoks, pH ve sıcaklık gradyanında bir sızıntı bölgesi hidrotermal höyüğündeki yapılandırılmış demir monosülfit çökeltilerinde evrimleştiğini" öne sürmüşlerdir. Fosilleşmiş sızıntı bölgesi metal sülfür çökeltilerinde gözlemlenen doğal olarak ortaya çıkan üç boyutlu bölmeler, bu inorganik bölmelerin serbest yaşayan prokaryotlarda bulunan hücre duvarlarının ve zarlarının öncüleri olduğunu göstermektedir. FeS ve NiS'in hidrotermal sıvının bileşenleri olan karbonmonoksit ve metilsülfürden asetil-metilsülfür sentezini katalize etme konusundaki bilinen kabiliyeti, biyotik öncesi sentezlerin bu metal-sülfür duvarlı bölmelerin iç yüzeylerinde meydana geldiğini göstermektedir."[193]
Bunlar, ultra-mafik olivinin deniz suyuyla serpantinleşmesi ve karbondioksit bakımından zengin okyanus suyuyla pH arayüzeyinin bir sonucu olarak hidrojen bakımından zengin sıvıların deniz tabanının altından çıktığı yerlerde oluşur. Bacalar, elektron vericilerin (moleküler hidrojen) elektron alıcılarla (karbondioksit) reaksiyona girdiği redoks reaksiyonlarından türetilen sürekli bir kimyasal enerji kaynağı oluşturur; bkz. demir-kükürt dünya teorisi. Bunlar ekzotermik reaksiyonlardır.[191]

Russell, alkali bacaların abiyogenez için ideal olan abiyojenik bir proton itici kuvvet kemiosmotik gradyanı yarattığını göstermiştir.[193] Mackinawite gibi demir-sülfür minerallerinden oluşan mikroskobik bölmeleri "organik molekülleri yoğunlaştırmak için doğal bir araç sağlar" ve bu mineral hücrelere Günter Wächtershäuser tarafından öngörülen katalitik özellikleri kazandırır.[194] İyonların membran boyunca bu hareketi iki faktörün kombinasyonuna bağlıdır:
- Konsantrasyon gradyanının neden olduğu difüzyon kuvveti - iyonlar dahil tüm partiküller yüksek konsantrasyondan düşük konsantrasyona doğru yayılma eğilimindedir.
- Elektriksel potansiyel gradyanının neden olduğu elektrostatik kuvvet - protonlar H+ gibi katyonlar elektriksel potansiyelden aşağı doğru, anyonlar ise ters yönde yayılma eğilimindedir.
Bu iki gradyan birlikte ele alındığında, abiyojenik sentez için enerji sağlayan bir elektrokimyasal gradyan olarak ifade edilebilir. Proton hareket kuvveti, bir membran boyunca proton ve voltaj gradyanlarının (proton konsantrasyonu ve elektrik potansiyelindeki farklılıklar) bir kombinasyonu olarak depolanan potansiyel enerjinin ölçüsü olarak tanımlanabilir.[143]
Derin okyanus hidrotermal bacalarındaki mineral parçacıklarının yüzeyleri enzimlerinkine benzer katalitik özelliklere sahiptir ve uygulanan bir voltajla veya H2 veya H2S ile reaksiyona girerek sudaki çözünmüş CO2'den metanol (CH3OH) ve formik, asetik ve pirüvik asitler gibi basit organik moleküller oluşturabilir.[195][196]
Martin tarafından 2016 yılında rapor edilen araştırma, yaşamın hidrotermal bacalarda ortaya çıktığı,[197][198] termodinamik dengesizlikteki kaya-su etkileşimleri tarafından yönlendirilen yerkabuğundaki spontane kimyanın yaşamın kökenini desteklediği ve arkea ve bakterilerin kurucu soylarının, enerji metabolizmasında terminal alıcı olarak CO2 kullanan H2'ye bağımlı ototroflar olduğu tezini desteklemektedir.[199][200][201] Martin, bu kanıtlara dayanarak, LUCA'nın "hayatta kalmak için havalandırmanın jeotermal enerjisine büyük ölçüde bağımlı olabileceğini" öne sürmektedir.[202] Derin deniz hidrotermal bacalarındaki gözeneklerin, biyokimyasal reaksiyonları teşvik eden membrana bağlı bölmeler tarafından işgal edildiği öne sürülmektedir.[203][204]
Kaplıcalar
[değiştir | kaynağı değiştir]Mulkidjanian ve ortak yazarlar, deniz ortamlarının hücrelerde evrensel olarak bulunan iyonik dengeyi ve bileşimi veya özellikle yüksek K+/Na+ oranı, Mn2+, Zn2+ ve fosfat konsantrasyonları açısından temel proteinler ve ribozimler için gerekli iyonları sağlamadığını düşünüyor. Dünya'da ihtiyaç duyulan koşulları taklit eden tek ortamın Kamçatka'dakine benzer sıcak su kaynakları olduğunu savunuyorlar.[205] Anoksik bir atmosfer altındaki bu ortamlardaki maden yatakları uygun pH değerine sahip olacak (oksijenli bir atmosferdeki mevcut havuzlar ise olmayacaktır), zararlı ultraviyole radyasyonu emen fotokatalitik sülfit minerallerinin çökeltilerini içerecek, substrat çözeltilerini hem hidrotermal ortamdaki kimyasal reaksiyonlar hem de bacalardan bitişik havuzlara taşınma sırasında biyomoleküllerin oluşumunu teşvik edecek UV ışığına maruz kalma yoluyla oluşturulan biyopolimerlerin kendiliğinden oluşumuna uygun konsantrasyonlara yoğunlaştıran ıslak-kuru döngülere sahip olacaktır.[206][207][208] Varsayılan biyotik öncesi ortamlar, LUCA'nın özelliklerini açıklamaya yardımcı olan ek bileşenlerle birlikte hidrotermal bacalara benzer.[155][205]
LUCA'ya ait olduğu düşünülen proteinlerin filogenomik ve jeokimyasal analizi, hücre içi sıvısının iyonik bileşiminin sıcak su kaynaklarında aynı olduğunu göstermektedir. LUCA muhtemelen büyümesi için sentezlenmiş organik maddeye bağımlıydı.[205] Deneyler, RNA benzeri polimerlerin çoklu ıslak-kuru döngülerde ve UV ışığına maruz bırakılarak sentezlenebileceğini göstermektedir. Bu polimerler yoğunlaşma sonrasında veziküller içinde kapsüllenmiş hale gelmiştir ki bu durum iyonik çözünen maddelerin yüksek konsantrasyonları nedeniyle tuzlu su koşullarında gerçekleşmez.[209] Kaplıcalarda biyomoleküllerin potansiyel kaynağı gezegenler arası toz parçacıkları, dünya dışı mermiler ya da atmosferik veya jeokimyasal sentezlerle taşınmasıdır. Kaplıca alanları Hadeen döneminde volkanik kara kütlelerinde bol miktarda bulunmuş olabilir.[155]
Kil
[değiştir | kaynağı değiştir]Kil hipotezi 1985 yılında Graham Cairns-Smith tarafından önerilmiştir.[210][211] Bu hipoteze göre karmaşık organik moleküller, sulu bir çözelti ile temas halindeki silikat kristallerinin önceden var olan, organik olmayan çoğalma yüzeylerinde kademeli olarak ortaya çıkmıştır. Kil minerali montmorillonitin, nükleotit monomerlerinden sulu çözeltide RNA polimerizasyonunu ve lipitlerden membran oluşumunu katalize ettiği gösterilmiştir.[212][213] 1998 yılında Hyman Hartman, "ilk organizmaların karbondioksiti oksalik asit ve diğer dikarboksilik asitlere bağlayan, kendi kendini kopyalayan demir açısından zengin killer olduğunu" öne sürmüştür. Bu çoğalan killer sistemi ve onların metabolik fenotipi daha sonra azotu bağlama yeteneği kazanarak kaplıcanın sülfür bakımından zengin bölgesine doğru evrimleşmiştir. Son olarak fosfat, nükleotitlerin ve fosfolipitlerin sentezine izin veren gelişen sisteme dahil edildi."[214]
Demir-kükürt dünyası
[değiştir | kaynağı değiştir]1980'lerde Günter Wächtershäuser ve Karl Popper, biyotik öncesi kimyasal yolların evrimi için demir-kükürt dünyası hipotezini ortaya atmıştır. Bu hipotez, günümüz biyokimyasını gazlardan organik yapı taşları sentezleyen ilkel reaksiyonlara kadar götürmektedir.[215][216] Wächtershäuser sistemleri yerleşik bir enerji kaynağına sahiptir: pirit gibi demir sülfitler. Bu metal sülfürlerin oksitlenmesiyle açığa çıkan enerji organik moleküllerin sentezini destekleyebilir. Bu tür sistemler, modern yaşam formlarından önce kendi kendini kopyalayan, metabolik olarak aktif varlıkları oluşturan otokatalitik setlere dönüşmüş olabilir.[217] 100°C'de sulu ortamda sülfürlerle yapılan deneyler küçük bir dipeptit verimi (%0,4 ile %12,4) ve daha küçük bir tripeptit verimi (%0,10) üretmiştir. Bununla birlikte, aynı koşullar altında dipeptitler hızla parçalanmıştır.[218]
Çeşitli modeller, RNA replikasyonunun daha sonra ortaya çıkmasına izin veren ilkel bir metabolizma varsaymaktadır. Krebs döngüsünün (sitrik asit döngüsü) aerobik organizmalarda enerji üretimindeki ve karmaşık organik kimyasalların biyosentezinde karbondioksit ve hidrojen iyonlarının çekilmesindeki merkezi önemi, bunun metabolizmanın evrimleşen ilk parçalarından biri olduğunu düşündürmektedir.[194] Buna paralel olarak jeokimyacı Jack W. Szostak ve Kate Adamala, ilkel protosellerde enzimatik olmayan RNA replikasyonunun yalnızca sitrik asit gibi zayıf katyon şelatörlerinin varlığında mümkün olduğunu göstermiştir. Bu, sitrik asidin ilkel metabolizmadaki merkezi rolüne dair daha fazla kanıt sağlamaktadır.[219] Russell "yaşamın amacının karbondioksiti hidrojenize etmek" olduğunu öne sürmüştür ("önce genetik" senaryosundan ziyade "önce metabolizma" senaryosunun bir parçası olarak).[217][220][221] Fizikçi Jeremy England, genel termodinamik değerlendirmelerden yola çıkarak yaşamın kaçınılmaz olduğunu savunmuştur.[222] Bu fikrin erken bir versiyonu Oparin'in 1924'te kendi kendini kopyalayan kesecikler için yaptığı öneriydi. 1980'lerde ve 1990'larda Wächtershäuser'in demir-kükürt dünyası teorisi ve Christian de Duve'nin tiyoester modelleri ortaya çıktı. Genler olmadan metabolizma için daha soyut ve teorik argümanlar arasında Freeman Dyson'ın matematiksel modeli ve Stuart Kauffman'ın 1980'lerdeki toplu otokatalitik kümeleri yer almaktadır. Kauffman'ın çalışması, hücrelerdeki biyokimyasal reaksiyonları yönlendirmede enerjinin rolünü göz ardı ettiği için eleştirilmiştir.[223]

Krebs döngüsü gibi çok adımlı bir biyokimyasal yol, bir mineral yüzeyinde kendiliğinden organize olmamıştır; öncesinde daha basit yolların olması gerekir.[224] Wood-Ljungdahl yolu, bir metal sülfit yüzeyinde kendi kendine örgütlenme ile uyumludur. Temel enzim birimi olan karbonmonoksit dehidrojenaz/asetil-CoA sentaz, reaksiyon merkezlerinde karışık nikel-demir-sülfür kümeleri içerir ve asetil-CoA oluşumunu katalize eder. Bununla birlikte, prebiyotik tiyolatlı ve tiyoester bileşiklerin hidrotermal bacaların varsayılan prebiyotik koşullarında birikmesi termodinamik ve kinetik olarak olası değildir.[225] Bir olasılık, sistein ve homosisteinin Strecker reaksiyonundan kaynaklanan nitrillerle reaksiyona girerek katalitik tiyol bakımından zengin polipeptitler oluşturmuş olabileceğidir.[226]
Çinko dünyası
[değiştir | kaynağı değiştir]Armen Mulkidjanian'ın çinko dünyası (Zn-dünyası) hipotezi, Wächtershäuser'in pirit hipotezini genişletmektedir.[227][228] Zn-dünyası teorisi, H2S bakımından zengin hidrotermal akışkanların soğuk ilkel okyanus (veya Darwin'in "sıcak küçük göleti") suyuyla etkileşime girerek metal sülfür parçacıklarını çökelttiğini öne sürer. Okyanus hidrotermal sistemleri, eski volkanojenik masif sülfit cevheri yataklarına yansıyan bölgesel bir yapıya sahiptir. Kilometrelerce çapa ulaşırlar ve Arkeen'e kadar uzanırlar. En bol bulunanlar pirit (FeS2), kalkopirit (CuFeS2) ve sfalerit (ZnS) olup bunlara galena (PbS) ve alabandit (MnS) de eklenir. ZnS ve MnS, örneğin ultraviyole ışıktan gelen radyasyon enerjisini depolamak için benzersiz bir yeteneğe sahiptir. Çoğalan moleküller ortaya çıkarken, ilkel atmosferik basınç Dünya yüzeyinin yakınında çökelecek kadar yüksekti (>100 bar) ve ultraviyole ışınımı şimdikinden 10 ila 100 kat daha yoğundu; dolayısıyla ZnS'nin aracılık ettiği fotosentetik özellikler, bilgi ve metabolik moleküllerin sentezi ve fotostabil nükleobazların seçimi için doğru enerji koşullarını sağlamıştır.[227][229]
Zn-dünyası teorisi, ilk protosellerin iç kısımlarının iyonik yapısına ilişkin kanıtlarla doldurulmuştur. Kanadalı biyokimyacı Archibald Macallum 1926'da kan ve lenf gibi vücut sıvılarının deniz suyuna benzerliğine dikkat çekmiştir;[230] ancak tüm hücrelerin inorganik bileşimi modern deniz suyundan farklıdır ve bu da Mulkidjanian ve meslektaşlarını jeokimyasal analiz ile modern hücrelerin inorganik iyon gereksinimlerinin filogenomik incelemesini birleştirerek ilk hücrelerin "kuluçkahanelerini" yeniden inşa etmeye yöneltmiştir. Yazarlar, her yerde bulunan ve çıkarım yoluyla ilkel olan proteinlerin ve işlevsel sistemlerin K+, Zn2+, Mn2+ ve [PO4]3-'e yakınlık ve işlevsel gereksinim gösterdiği sonucuna varmıştır. Jeokimyasal yeniden yapılandırma, bu iyonik bileşimin okyanusta var olamayacağını, ancak iç jeotermal sistemlerle uyumlu olduğunu göstermektedir. Oksijeni tükenmiş, CO2 ağırlıklı ilkel atmosferde, jeotermal alanların yakınındaki su yoğuşmalarının kimyası modern hücrelerin iç ortamına benzeyecektir. Bu nedenle, hücre öncesi evrim, metal sülfürlerle karışık gözenekli silikat mineralleriyle kaplı ve K+, Zn2+ ve fosfor bileşikleri bakımından zengin sığ "Darwin havuzlarında" gerçekleşmiş olabilir.[231][232]
Homokiralite
[değiştir | kaynağı değiştir]
Homokiralite, kiral (ayna simetrisi olmayan) birimlerden oluşan malzemelerin geometrik tekdüzeliğidir. Canlı organizmalar aynı kiraliteye (ellilik) sahip molekülleri kullanırlar: neredeyse istisnasız[234] olarak amino asitler sol elli iken nükleotitler ve şekerler sağ ellidir. Kiral moleküller sentezlenebilir, ancak kiral bir kaynak veya kiral bir katalizör olmadığında, her iki formun 50/50 (rasemik) karışımı halinde oluşurlar. Rasemik başlangıç malzemelerinden rasemik olmayan karışımların üretimi için bilinen mekanizmalar şunları içerir: elektrozayıf etkileşim gibi asimetrik fiziksel yasalar; dairesel polarize ışık, kuvars kristalleri veya Dünya'nın dönüşünden kaynaklananlar gibi asimetrik ortamlar, rasemik sentez sırasında istatistiksel dalgalanmalar[233] ve kendiliğinden simetri kırılması.[235][236][237] Bir kez oluşturulduktan sonra, kiralite için seçim yapılacaktır.[238] Popülasyondaki küçük bir sapma (enantiyomerik fazlalık), Soai reaksiyonunda olduğu gibi asimetrik otokataliz ile büyük bir sapmaya dönüştürülebilir.[239] Asimetrik otokatalizde katalizör kiral bir moleküldür, bu da kiral bir molekülün kendi üretimini katalizlediği anlamına gelir. Polarize ışıkla üretilebileceği gibi başlangıçtaki bir enantiyomerik fazlalık, daha sonra daha bol olan enantiyomerin diğerine üstün gelmesini sağlar.[240] Murchison meteorunda L-alanin amino asidi D formundan iki kat daha sık ve L-glutamik asit D muadilinden üç kat daha fazla olduğu için homokiralite uzayda başlamış olabilir.[241][242] Meteoritlerden elde edilen amino asitler sol-elli bir eğilim gösterirken, şekerler canlı organizmalarda olduğu gibi ağırlıklı olarak sağ-elli bir eğilim göstermekte ve bu da bu bileşiklerin abiyojenik bir kökene sahip olduğunu düşündürmektedir.[243]
Ayrıca bakınız
[değiştir | kaynağı değiştir]Notlar
[değiştir | kaynağı değiştir]- ^ Reaksiyonlar şunlardır:
- FeS + H2S → FeS2 + 2H+ + 2e−
- FeS + H2S + CO2 → FeS2 + HCOOH
Kaynakça
[değiştir | kaynağı değiştir]- ^ a b Walker, Sara I.; Packard, N.; Cody, G. D. (13 Kasım 2017). "Re-conceptualizing the origins of life". Philosophical Transactions of the Royal Society A. 375 (2109): 20160337. Bibcode:2017RSPTA.37560337W. doi:10.1098/rsta.2016.0337. PMC 5686397 $2. PMID 29133439.
- ^ a b c "NASA Astrobiology Strategy" (PDF). NASA. 2015. 22 Aralık 2016 tarihinde kaynağından (PDF) arşivlendi. Erişim tarihi: 24 Eylül 2017.
- ^ Trifonov, Edward N. (17 Mart 2011). "Vocabulary of Definitions of Life Suggests a Definition". Journal of Biomolecular Structure and Dynamics. 29 (2): 259-266. doi:10.1080/073911011010524992. PMID 21875147.
- ^ Voytek, Mary A. (6 Mart 2021). "About Life Detection". NASA. 16 Ağustos 2021 tarihinde kaynağından arşivlendi. Erişim tarihi: 8 Mart 2021.
- ^ a b Witzany, Guenther (2016). "Crucial steps to life: From chemical reactions to code using agents" (PDF). BioSystems. 140: 49-57. doi:10.1016/j.biosystems.2015.12.007. PMID 26723230. 31 Ekim 2018 tarihinde kaynağından arşivlendi (PDF). Erişim tarihi: 2 Nisan 2023.
- ^ a b Howell, Elizabeth (8 Aralık 2014). "How Did Life Become Complex, And Could It Happen Beyond Earth?". Astrobiology Magazine. Archived from the original on 15 Şubat 2018. Erişim tarihi: 14 Nisan 2022.
- ^ Oparin, Aleksandr Ivanovich (2003) [1938]. The Origin of Life. Morgulis, Sergius tarafından çevrildi (2 bas.). Mineola, New York: Courier. ISBN 978-0486495224. 2 Nisan 2023 tarihinde kaynağından arşivlendi. Erişim tarihi: 2 Nisan 2023.
- ^ a b Peretó, Juli (2005). "Controversies on the origin of life" (PDF). International Microbiology. 8 (1): 23-31. PMID 15906258. 24 Ağustos 2015 tarihinde kaynağından (PDF) arşivlendi. Erişim tarihi: 1 Haziran 2015.
- ^ Compare: Scharf, Caleb (18 Aralık 2015). "A Strategy for Origins of Life Research". Astrobiology. 15 (12): 1031-1042. Bibcode:2015AsBio..15.1031S. doi:10.1089/ast.2015.1113. PMC 4683543 $2. PMID 26684503.
What do we mean by the origins of life (OoL)? ... Since the early 20th century the phrase OoL has been used to refer to the events that occurred during the transition from non-living to living systems on Earth, i.e., the origin of terrestrial biology (Oparin, 1924; Haldane, 1929). The term has largely replaced earlier concepts such as abiogenesis (Kamminga, 1980; Fry, 2000).
- ^ a b Weiss, M. C.; Sousa, F. L.; Mrnjavac, N.; Neukirchen, S.; Roettger, M.; Nelson-Sathi, S.; Martin, W.F. (2016). "The physiology and habitat of the last universal common ancestor" (PDF). Nature Microbiology. 1 (9): 16116. doi:10.1038/NMICROBIOL.2016.116. PMID 27562259. 29 Ocak 2023 tarihinde kaynağından arşivlendi (PDF). Erişim tarihi: 2 Nisan 2023.
- ^ Tirard, Stephane (20 Nisan 2015). Abiogenesis – Definition. Encyclopedia of Astrobiology. s. 1. doi:10.1007/978-3-642-27833-4_2-4. ISBN 978-3-642-27833-4.
Thomas Huxley (1825–1895) used the term abiogenesis in an important text published in 1870. He strictly made the difference between spontaneous generation, which he did not accept, and the possibility of the evolution of matter from inert to living, without any influence of life. ... Since the end of the nineteenth century, evolutive abiogenesis means increasing complexity and evolution of matter from inert to living state in the abiotic context of evolution of primitive Earth.
- ^ Luisi, Pier Luigi (2018). The Emergence of Life: From Chemical Origins to Synthetic Biology. Cambridge University Press. s. 416. ISBN 9781108735506.
However, the turning point of non-life to life has never been put into one experimental set up. There are, of course, several hypotheses, and this plethora of ideas means already that we do not have a convincing one.
- ^ Graham, Robert W. (February 1990). "Extraterrestrial Life in the Universe" (PDF). NASA (NASA Technical Memorandum 102363). Lewis Research Center, Cleveland, Ohio. 3 Eylül 2014 tarihinde kaynağından arşivlendi (PDF). Erişim tarihi: 2 Haziran 2015.
- ^ Altermann 2009, s. xvii
- ^ Oparin 1953, s. vi
- ^ Warmflash, David; Warmflash, Benjamin (November 2005). "Did Life Come from Another World?". Scientific American. 293 (5): 64-71. Bibcode:2005SciAm.293e..64W. doi:10.1038/scientificamerican1105-64. PMID 16318028.
- ^ Yarus 2010, s. 47
- ^ Ward, Peter; Kirschvink, Joe (2015). A New History of Life: the radical discoveries about the origins and evolution of life on earth. Bloomsbury Press. ss. 39-40. ISBN 978-1608199105.
- ^ Sheldon 2005
- ^ Lennox 2001, ss. 229–258
- ^ a b Bernal 1967
- ^ Balme, D. M. (1962). "Development of Biology in Aristotle and Theophrastus: Theory of Spontaneous Generation". Phronesis. 7 (1–2): 91-104. doi:10.1163/156852862X00052.
- ^ Ross 1652
- ^ Dobell 1960
- ^ Bondeson 1999
- ^ Levine, R.; Evers, C. "The Slow Death of Spontaneous Generation (1668-1859)". 26 Nisan 2008 tarihinde kaynağından arşivlendi. Erişim tarihi: 18 Nisan 2013.
- ^ Oparin 1953, s. 196
- ^ Tyndall 1905, IV, XII (1876), XIII (1878)
- ^ Horneck, Gerda; Klaus, David M.; Mancinelli, Rocco L. (March 2010). "Space Microbiology". Microbiology and Molecular Biology Reviews. 74 (1): 121-156. Bibcode:2010MMBR...74..121H. doi:10.1128/MMBR.00016-09. PMC 2832349 $2. PMID 20197502.
- ^ Wickramasinghe, Chandra (2011). "Bacterial morphologies supporting cometary panspermia: a reappraisal". International Journal of Astrobiology. 10 (1): 25-30. Bibcode:2011IJAsB..10...25W. CiteSeerX 10.1.1.368.4449 $2. doi:10.1017/S1473550410000157.
- ^ Rampelotto, P. H. (2010). "Panspermia: A promising field of research". In: Astrobiology Science Conference. Abs 5224.
- ^ Chang, Kenneth (12 Eylül 2016). "Visions of Life on Mars in Earth's Depths". The New York Times. 12 Eylül 2016 tarihinde kaynağından arşivlendi. Erişim tarihi: 12 Eylül 2016.
- ^ "Letter no. 7471, Charles Darwin to Joseph Dalton Hooker, 1 February (1871)". Darwin Correspondence Project. 7 Temmuz 2020 tarihinde kaynağından arşivlendi. Erişim tarihi: 7 Temmuz 2020.
- ^ Priscu, John C. "Origin and Evolution of Life on a Frozen Earth". Arlington County, Virginia: National Science Foundation. 18 Aralık 2013 tarihinde kaynağından arşivlendi. Erişim tarihi: 1 Mart 2014.
- ^ Marshall, Michael (11 Kasım 2020). "Charles Darwin's hunch about early life was probably right". BBC News. 11 Kasım 2020 tarihinde kaynağından arşivlendi. Erişim tarihi: 11 Kasım 2020.
- ^ Bahadur, Krishna (1973). "Photochemical Formation of Self–sustaining Coacervates" (PDF). Proceedings of the Indian National Science Academy. 39 (4): 455-467. doi:10.1016/S0044-4057(75)80076-1. PMID 1242552. 19 Ekim 2013 tarihinde kaynağından (PDF) arşivlendi.
- ^ Bahadur, Krishna (1975). "Photochemical Formation of Self-Sustaining Coacervates". Zentralblatt für Bakteriologie, Parasitenkunde, Infektionskrankheiten und Hygiene (Central Journal for Bacteriology, Parasitology, Infectious Diseases and Hygiene). 130 (3): 211-218. doi:10.1016/S0044-4057(75)80076-1. OCLC 641018092. PMID 1242552. 13 Aralık 2022 tarihinde kaynağından arşivlendi. Erişim tarihi: 2 Nisan 2023.
- ^ Bryson 2004, ss. 300–302
- ^ Bernal 1951
- ^ Martin, William F. (January 2003). "On the origins of cells: a hypothesis for the evolutionary transitions from abiotic geochemistry to chemoautotrophic prokaryotes, and from prokaryotes to nucleated cells". Phil. Trans. R. Soc. Lond. A. 358 (1429): 59-83. doi:10.1098/rstb.2002.1183. PMC 1693102 $2. PMID 12594918.
- ^ Bernal, John Desmond (September 1949). "The Physical Basis of Life". Proceedings of the Physical Society, Section A. 62 (9): 537-558. Bibcode:1949PPSA...62..537B. doi:10.1088/0370-1298/62/9/301.
- ^ Miller, Stanley L. (15 Mayıs 1953). "A Production of Amino Acids Under Possible Primitive Earth Conditions". Science. 117 (3046): 528-529. Bibcode:1953Sci...117..528M. doi:10.1126/science.117.3046.528. PMID 13056598.
- ^ Parker, Eric T.; Cleaves, Henderson J.; Dworkin, Jason P.; Glavin, Daniel P.; Callahan, Michael; Aubrey, Andrew; Lazcano, Antonio; Bada, Jeffrey L. (5 Nisan 2011). "Primordial synthesis of amines and amino acids in a 1958 Miller H2S-rich spark discharge experiment". PNAS. 108 (14): 5526-5531. Bibcode:2011PNAS..108.5526P. doi:10.1073/pnas.1019191108. PMC 3078417 $2. PMID 21422282.
- ^ Bernal 1967, s. 143
- ^ a b Cleaves, H. James; Chalmers, John H.; Lazcano, Antonio; Miller, Stanley L.; Bada, Jeffrey L. (April 2008). "A Reassessment of Prebiotic Organic Synthesis in Neutral Planetary Atmospheres". Origins of Life and Evolution of Biospheres. 38 (2): 105-115. Bibcode:2008OLEB...38..105C. doi:10.1007/s11084-007-9120-3. PMID 18204914.
- ^ Chyba, Christopher F. (13 Mayıs 2005). "Rethinking Earth's Early Atmosphere". Science. 308 (5724): 962-963. doi:10.1126/science.1113157. PMID 15890865.
- ^ Barton et al. 2007, ss. 93–95
- ^ Bada & Lazcano 2009, ss. 56–57
- ^ Bada, Jeffrey L.; Lazcano, Antonio (2 Mayıs 2003). "Prebiotic Soup – Revisiting the Miller Experiment" (PDF). Science. 300 (5620): 745-746. doi:10.1126/science.1085145. PMID 12730584. 4 Mart 2016 tarihinde kaynağından arşivlendi (PDF). Erişim tarihi: 13 Haziran 2015.
- ^ Marigo, Paola (6 Temmuz 2020). "Carbon star formation as seen through the non-monotonic initial–final mass relation". Nature Astronomy. 152 (11): 1102-1110. arXiv:2007.04163 $2. Bibcode:2020NatAs...4.1102M. doi:10.1038/s41550-020-1132-1. 16 Şubat 2023 tarihinde kaynağından arşivlendi. Erişim tarihi: 2 Nisan 2023.
- ^ Marigo, Paola (6 Temmuz 2020). "Carbon star formation as seen through the non-monotonic initial–final mass relation". Nature Astronomy. 152 (11): 1102-1110. arXiv:2007.04163 $2. Bibcode:2020NatAs...4.1102M. doi:10.1038/s41550-020-1132-1. 16 Şubat 2023 tarihinde kaynağından arşivlendi. Erişim tarihi: 7 Temmuz 2020.
- ^ "WMAP- Life in the Universe". wmap.gsfc.nasa.gov. 29 Ocak 2023 tarihinde kaynağından arşivlendi. Erişim tarihi: 26 Mart 2023.
- ^ "Formation of Solar Systems: Solar Nebular Theory". University of Massachusetts Amherst. 27 Eylül 2019 tarihinde kaynağından arşivlendi. Erişim tarihi: 27 Eylül 2019.
- ^ "Age of the Earth". United States Geological Survey. 9 Temmuz 2007. 23 Aralık 2005 tarihinde kaynağından arşivlendi. Erişim tarihi: 10 Ocak 2006.
- ^ Dalrymple 2001, ss. 205–221
- ^ Fesenkov 1959, s. 9
- ^ Bottke, W. F.; Vokrouhlický, D.; Marchi, S.; Swindle, T.; Scott, E. R. D.; Weirich, J. R.; Levison, H. (17 Nisan 2015). "Dating the Moon-forming impact event with asteroidal meteorites". Science. 348 (6232): 321-323. Bibcode:2015Sci...348..321B. doi:10.1126/science.aaa0602
 .
. - ^ Kasting, James F. (12 Şubat 1993). "Earth's Early Atmosphere" (PDF). Science. 259 (5097): 920-926. Bibcode:1993Sci...259..920K. doi:10.1126/science.11536547. PMID 11536547. 10 Ekim 2015 tarihinde kaynağından (PDF) arşivlendi. Erişim tarihi: 28 Temmuz 2015.
- ^ a b c d e f g Follmann, Hartmut; Brownson, Carol (November 2009). "Darwin's warm little pond revisited: from molecules to the origin of life". Naturwissenschaften. 96 (11): 1265-1292. Bibcode:2009NW.....96.1265F. doi:10.1007/s00114-009-0602-1. PMID 19760276.
- ^ Morse, John (September 1998). "Hadean Ocean Carbonate Geochemistry". Aquatic Geochemistry. 4 (3/4): 301-319. Bibcode:1998MinM...62.1027M. doi:10.1023/A:1009632230875.
- ^ Morse, John W.; MacKenzie, Fred T. (1998). "Hadean Ocean Carbonate Geochemistry". Aquatic Geochemistry. 4 (3–4): 301-319. Bibcode:1998MinM...62.1027M. doi:10.1023/A:1009632230875.
- ^ Wilde, Simon A.; Valley, John W.; Peck, William H.; Graham, Colin M. (11 Ocak 2001). "Evidence from detrital zircons for the existence of continental crust and oceans on the Earth 4.4 Gyr ago" (PDF). Nature. 409 (6817): 175-178. Bibcode:2001Natur.409..175W. doi:10.1038/35051550. PMID 11196637. 5 Haziran 2015 tarihinde kaynağından arşivlendi (PDF). Erişim tarihi: 3 Haziran 2015.
- ^ Rosing, Minik T.; Bird, Dennis K.; Sleep, Norman H.; Glassley, William; Albarède, Francis (22 Mart 2006). "The rise of continents – An essay on the geologic consequences of photosynthesis". Palaeogeography, Palaeoclimatology, Palaeoecology. 232 (2–4): 99-113. Bibcode:2006PPP...232...99R. doi:10.1016/j.palaeo.2006.01.007. 14 Temmuz 2015 tarihinde kaynağından arşivlendi (PDF). Erişim tarihi: 8 Haziran 2015.
- ^ a b c Dodd, Matthew S.; Papineau, Dominic; Grenne, Tor; Slack, John F.; Rittner, Martin; Pirajno, Franco; O'Neil, Jonathan; Little, Crispin T.S. (1 Mart 2017). "Evidence for early life in Earth's oldest hydrothermal vent precipitates". Nature. 543 (7643): 60-64. Bibcode:2017Natur.543...60D. doi:10.1038/nature21377. PMID 28252057. 8 Eylül 2017 tarihinde kaynağından arşivlendi. Erişim tarihi: 2 Mart 2017.
- ^ Gomes, Rodney; Levison, Hal F.; Tsiganis, Kleomenis; Morbidelli, Alessandro (26 Mayıs 2005). "Origin of the cataclysmic Late Heavy Bombardment period of the terrestrial planets". Nature. 435 (7041): 466-469. Bibcode:2005Natur.435..466G. doi:10.1038/nature03676. PMID 15917802.
- ^ Sleep, Norman H.; Zahnle, Kevin J.; Kasting, James F.; Morowitz, Harold J. (9 Kasım 1989). "Annihilation of ecosystems by large asteroid impacts on early Earth". Nature. 342 (6246): 139-142. Bibcode:1989Natur.342..139S. doi:10.1038/342139a0. PMID 11536616.
- ^ Chyba, Christopher; Sagan, Carl (9 Ocak 1992). "Endogenous production, exogenous delivery and impact-shock synthesis of organic molecules: an inventory for the origins of life". Nature. 355 (6356): 125-132. Bibcode:1992Natur.355..125C. doi:10.1038/355125a0. PMID 11538392.
- ^ Furukawa, Yoshihiro; Sekine, Toshimori; Oba, Masahiro; Kakegawa, Takeshi; Nakazawa, Hiromoto (January 2009). "Biomolecule formation by oceanic impacts on early Earth". Nature Geoscience. 2 (1): 62-66. Bibcode:2009NatGe...2...62F. doi:10.1038/NGEO383.
- ^ Maher, Kevin A.; Stevenson, David J. (18 Şubat 1988). "Impact frustration of the origin of life". Nature. 331 (6157): 612-614. Bibcode:1988Natur.331..612M. doi:10.1038/331612a0. PMID 11536595.
- ^ Mann, Adam (24 Ocak 2018). "Bashing holes in the tale of Earth's troubled youth". Nature. 553 (7689): 393-395. Bibcode:2018Natur.553..393M. doi:10.1038/d41586-018-01074-6.
- ^ Davies 1999, s. 155
- ^ Bock & Goode 1996
- ^ Schopf, J. William; Kudryavtsev, Anatoliy B.; Czaja, Andrew D.; Tripathi, Abhishek B. (5 Ekim 2007). "Evidence of Archean life: Stromatolites and microfossils". Precambrian Research. 158 (3–4): 141-155. Bibcode:2007PreR..158..141S. doi:10.1016/j.precamres.2007.04.009.
- ^ Schopf, J. William (29 Haziran 2006). "Fossil evidence of Archaean life". Philosophical Transactions of the Royal Society B. 361 (1470): 869-885. doi:10.1098/rstb.2006.1834. PMC 1578735 $2. PMID 16754604.
- ^ Raven & Johnson 2002, s. 68
- ^ Djokic, Tara; Van Kranendonk, Martin J.; Campbell, Kathleen A.; Walter, Malcolm R.; Ward, Colin R. (9 Mayıs 2017). "Earliest signs of life on land preserved in ca. 3.5 Gao hot spring deposits". Nature Communications. 8: 15263. Bibcode:2017NatCo...815263D. doi:10.1038/ncomms15263. PMC 5436104 $2. PMID 28486437.
- ^ Schopf, J. William; Kitajima, Kouki; Spicuzza, Michael J.; Kudryavtsev, Anatolly B.; Valley, John W. (2017). "SIMS analyses of the oldest known assemblage of microfossils document their taxon-correlated carbon isotope compositions". PNAS. 115 (1): 53-58. Bibcode:2018PNAS..115...53S. doi:10.1073/pnas.1718063115. PMC 5776830 $2. PMID 29255053.
- ^ Tyrell, Kelly April (18 Aralık 2017). "Oldest fossils ever found show life on Earth began before 3.5 billion years ago". University of Wisconsin-Madison. 10 Şubat 2021 tarihinde kaynağından arşivlendi. Erişim tarihi: 18 Aralık 2017.
- ^ Ohtomo, Yoko; Kakegawa, Takeshi; Ishida, Akizumi; Nagase, Toshiro; Rosing, Minik T. (January 2014). "Evidence for biogenic graphite in early Archaean Isua metasedimentary rocks". Nature Geoscience. 7 (1): 25-28. Bibcode:2014NatGe...7...25O. doi:10.1038/ngeo2025.
- ^ Noffke, Nora; Christian, Daniel; Wacey, David; Hazen, Robert M. (16 Kasım 2013). "Microbially Induced Sedimentary Structures Recording an Ancient Ecosystem in the ca. 3.48 Gyo Dresser Formation, Pilbara, Western Australia". Astrobiology. 13 (12): 1103-1124. Bibcode:2013AsBio..13.1103N. doi:10.1089/ast.2013.1030. PMC 3870916 $2. PMID 24205812.
- ^ Davies 1999
- ^ Hassenkam, T.; Andersson, M. P.; Dalby, K. N.; Mackenzie, D.M.A.; Rosing, M.T. (2017). "Elements of Eoarchean life trapped in mineral inclusions". Nature. 548 (7665): 78-81. Bibcode:2017Natur.548...78H. doi:10.1038/nature23261. PMID 28738409.
- ^ O'Donoghue, James (21 Ağustos 2011). "Oldest reliable fossils show early life was a beach". New Scientist. 211: 13. doi:10.1016/S0262-4079(11)62064-2. 30 Haziran 2015 tarihinde kaynağından arşivlendi.
- ^ Wacey, David; Kilburn, Matt R.; Saunders, Martin; Cliff, John; Brasier, Martin D. (October 2011). "Microfossils of sulphur-metabolizing cells in 3.4-billion-year-old rocks of Western Australia". Nature Geoscience. 4 (10): 698-702. Bibcode:2011NatGe...4..698W. doi:10.1038/ngeo1238.
- ^ Bell, Elizabeth A.; Boehnike, Patrick; Harrison, T. Mark; Mao, Wendy L. (19 Ekim 2015). "Potentially biogenic carbon preserved in a 4.1 billion-year-old zircon". PNAS. 112 (47): 14518-14521. Bibcode:2015PNAS..11214518B. doi:10.1073/pnas.1517557112. PMC 4664351 $2. PMID 26483481. Early edition, published online before print.
- ^ Baumgartner, Rafael; Van Kranendonk, Martin; Wacey, David; Fiorentini, Marco; Saunders, Martin; Caruso, Caruso; Pages, Anais; Homann, Martin; Guagliardo, Paul (2019). "Nano−porous pyrite and organic matter in 3.5-billion-year-old stromatolites record primordial life" (PDF). Geology. 47 (11): 1039-1043. Bibcode:2019Geo....47.1039B. doi:10.1130/G46365.1. 5 Aralık 2020 tarihinde kaynağından arşivlendi (PDF). Erişim tarihi: 2 Nisan 2023.
- ^ Djokic, Tara; Van Kranendonk, Martin; Cambell, Kathleen; Walter, Malcolm (2017). "Earliest signs of life on land preserved in ca. 3.5 Ga hot spring deposits". Nature Communications. 3.
- ^ Landau, Elizabeth (12 Ekim 2016). "Building Blocks of Life's Building Blocks Come From Starlight". NASA. 13 Ekim 2016 tarihinde kaynağından arşivlendi. Erişim tarihi: 13 Ekim 2016.
- ^ a b Ehrenfreund, Pascale; Cami, Jan (December 2010). "Cosmic carbon chemistry: from the interstellar medium to the early Earth". Cold Spring Harbor Perspectives in Biology. 2 (12): a002097. doi:10.1101/cshperspect.a002097. PMC 2982172 $2. PMID 20554702.
- ^ Geballe, Thomas R.; Najarro, Francisco; Figer, Donald F.; Schlegelmilch, Barret W.; de la Fuente, Diego (10 Kasım 2011). "Infrared diffuse interstellar bands in the Galactic Centre region". Nature. 479 (7372): 200-202. arXiv:1111.0613 $2. Bibcode:2011Natur.479..200G. doi:10.1038/nature10527. PMID 22048316.
- ^ Klyce 2001
- ^ a b c d Hoover, Rachel (21 Şubat 2014). "Need to Track Organic Nano-Particles Across the Universe? NASA's Got an App for That". Ames Research Center. NASA. 6 Eylül 2015 tarihinde kaynağından arşivlendi. Erişim tarihi: 22 Haziran 2015.
- ^ Goncharuk, Vladislav V.; Zui, O. V. (February 2015). "Water and carbon dioxide as the main precursors of organic matter on Earth and in space". Journal of Water Chemistry and Technology. 37 (1): 2-3. doi:10.3103/S1063455X15010026.
- ^ Abou Mrad, Ninette; Vinogradoff, Vassilissa; Duvernay, Fabrice; Danger, Grégoire; Theulé, Patrice; Borget, Fabien; Chiavassa, Thierry (2015). "Laboratory experimental simulations: Chemical evolution of the organic matter from interstellar and cometary ice analogs". Bulletin de la Société Royale des Sciences de Liège. 84: 21-32. Bibcode:2015BSRSL..84...21A. 13 Nisan 2015 tarihinde kaynağından arşivlendi. Erişim tarihi: 6 Nisan 2015.
- ^ Oba, Yasuhiro (26 Nisan 2022). "Identifying the wide diversity of extraterrestrial purine and pyrimidine nucleobases in carbonaceous meteorites". Nature Communications. 13 (2008): 2008. Bibcode:2022NatCo..13.2008O. doi:10.1038/s41467-022-29612-x. PMC 9042847 $2. PMID 35473908.
- ^ "'Life chemical' detected in comet". BBC News. Londra. 18 Ağustos 2009. 25 Mayıs 2015 tarihinde kaynağından arşivlendi. Erişim tarihi: 23 Haziran 2015.
- ^ Thompson, William Reid; Murray, B. G.; Khare, Bishun Narain; Sagan, Carl (30 Aralık 1987). "Coloration and darkening of methane clathrate and other ices by charged particle irradiation: Applications to the outer solar system". Journal of Geophysical Research. 92 (A13): 14933-14947. Bibcode:1987JGR....9214933T. doi:10.1029/JA092iA13p14933. PMID 11542127.
- ^ Goldman, Nir; Tamblyn, Isaac (20 Haziran 2013). "Prebiotic Chemistry within a Simple Impacting Icy Mixture". Journal of Physical Chemistry A. 117 (24): 5124-5131. Bibcode:2013JPCA..117.5124G. doi:10.1021/jp402976n. PMID 23639050. 21 Temmuz 2018 tarihinde kaynağından arşivlendi. Erişim tarihi: 2 Nisan 2023.
- ^ "NASA Ames PAH IR Spectroscopic Database". NASA. 29 Haziran 2015 tarihinde kaynağından arşivlendi. Erişim tarihi: 17 Haziran 2015.
- ^ a b c Hudgins, Douglas M.; Bauschlicher, Charles W. Jr.; Allamandola, Louis J. (10 Ekim 2005). "Variations in the Peak Position of the 6.2 μm Interstellar Emission Feature: A Tracer of N in the Interstellar Polycyclic Aromatic Hydrocarbon Population". The Astrophysical Journal. 632 (1): 316-332. Bibcode:2005ApJ...632..316H. CiteSeerX 10.1.1.218.8786 $2. doi:10.1086/432495.
- ^ a b c Des Marais, David J.; Allamandola, Louis J.; Sandford, Scott; Mattioda, Andrew; Gudipati, Murthy; Roser, Joseph; Bramall, Nathan; Nuevo, Michel; Boersma, Christiaan; Bernstein, Max; Peeters, Els; Cami, Jan; Cook, Jamie Elsila; Dworkin, Jason (2009). "Cosmic Distribution of Chemical Complexity". Ames Research Center. Mountain View, California: NASA. 27 Şubat 2014 tarihinde kaynağından arşivlendi. Erişim tarihi: 24 Haziran 2015.
- ^ a b Carey, Bjorn (18 Ekim 2005). "Life's Building Blocks 'Abundant in Space'". Space.com. Watsonville, California: Imaginova. 26 Haziran 2015 tarihinde kaynağından arşivlendi. Erişim tarihi: 23 Haziran 2015.
- ^ García-Hernández, Domingo. A.; Manchado, Arturo; García-Lario, Pedro; Stanghellini, Letizia; Villaver, Eva; Shaw, Richard A.; Szczerba, Ryszard; Perea-Calderón, Jose Vicente (20 Kasım 2010). "Formation of Fullerenes in H-Containing Planetary Nebulae". The Astrophysical Journal Letters. 724 (1): L39-L43. arXiv:1009.4357 $2. Bibcode:2010ApJ...724L..39G. doi:10.1088/2041-8205/724/1/L39.
- ^ d'Ischia, Marco; Manini, Paola; Moracci, Marco; Saladino, Raffaele; Ball, Vincent; Thissen, Helmut; Evans, Richard A.; Puzzarini, Cristina; Barone, Vincenzo (21 Ağustos 2019). "Astrochemistry and Astrobiology: Materials Science in Wonderland?". International Journal of Molecular Sciences. 20 (17): 4079. doi:10.3390/ijms20174079. PMC 6747172 $2. PMID 31438518.
- ^ Gudipati, Murthy S.; Yang, Rui (1 Eylül 2012). "In-situ Probing of Radiation-induced Processing of Organics in Astrophysical Ice Analogs – Novel Laser Desorption Laser Ionization Time-of-flight Mass Spectroscopic Studies". The Astrophysical Journal Letters. 756 (1): L24. Bibcode:2012ApJ...756L..24G. doi:10.1088/2041-8205/756/1/L24.
- ^ a b Gallori, Enzo (June 2011). "Astrochemistry and the origin of genetic material". Rendiconti Lincei. 22 (2): 113-118. doi:10.1007/s12210-011-0118-4. "Paper presented at the Symposium 'Astrochemistry: molecules in space and time' (Rome, 4–5 November 2010), sponsored by Fondazione 'Guido Donegani', Accademia Nazionale dei Lincei."
- ^ Martins, Zita (February 2011). "Organic Chemistry of Carbonaceous Meteorites". Elements. 7 (1): 35-40. doi:10.2113/gselements.7.1.35.
- ^ Martins, Zita; Botta, Oliver; Fogel, Marilyn L.; Sephton, Mark A.; Glavin, Daniel P.; Watson, Jonathan S.; Dworkin, Jason P.; Schwartz, Alan W.; Ehrenfreund, Pascale (15 Haziran 2008). "Extraterrestrial nucleobases in the Murchison meteorite". Earth and Planetary Science Letters. 270 (1–2): 130-136. arXiv:0806.2286 $2. Bibcode:2008E&PSL.270..130M. doi:10.1016/j.epsl.2008.03.026.
- ^ Callahan, Michael P.; Smith, Karen E.; Cleaves, H. James, II; Ruzica, Josef; Stern, Jennifer C.; Glavin, Daniel P.; House, Christopher H.; Dworkin, Jason P. (23 Ağustos 2011). "Carbonaceous meteorites contain a wide range of extraterrestrial nucleobases". PNAS. 108 (34): 13995-13998. Bibcode:2011PNAS..10813995C. doi:10.1073/pnas.1106493108. PMC 3161613 $2. PMID 21836052.
- ^ Steigerwald, John (8 Ağustos 2011). "NASA Researchers: DNA Building Blocks Can Be Made in Space". Goddard Space Flight Center. NASA. 23 Haziran 2015 tarihinde kaynağından arşivlendi. Erişim tarihi: 23 Haziran 2015.
- ^ Kwok, Sun; Zhang, Yong (3 Kasım 2011). "Mixed aromatic–aliphatic organic nanoparticles as carriers of unidentified infrared emission features". Nature. 479 (7371): 80-83. Bibcode:2011Natur.479...80K. doi:10.1038/nature10542. PMID 22031328.
- ^ Jørgensen, Jes K.; Favre, Cécile; Bisschop, Suzanne E.; Bourke, Tyler L.; van Dishoeck, Ewine F.; Schmalzl, Markus (2012). "Detection of the simplest sugar, glycolaldehyde, in a solar-type protostar with ALMA" (PDF). The Astrophysical Journal Letters. 757 (1): L4. arXiv:1208.5498 $2. Bibcode:2012ApJ...757L...4J. doi:10.1088/2041-8205/757/1/L4. 24 Eylül 2015 tarihinde kaynağından arşivlendi (PDF). Erişim tarihi: 23 Haziran 2015.
- ^ Furukawa, Yoshihiro; Chikaraishi, Yoshito; Ohkouchi, Naohiko; Ogawa, Nanako O.; Glavin, Daniel P.; Dworkin, Jason P.; Abe, Chiaki; Nakamura, Tomoki (13 Kasım 2019). "Extraterrestrial ribose and other sugars in primitive meteorites". PNAS. 116 (49): 24440-24445. Bibcode:2019PNAS..11624440F. doi:10.1073/pnas.1907169116. PMC 6900709 $2. PMID 31740594.
- ^ Oró, Joan; Kimball, Aubrey P. (February 1962). "Synthesis of purines under possible primitive earth conditions: II. Purine intermediates from hydrogen cyanide". Archives of Biochemistry and Biophysics. 96 (2): 293-313. doi:10.1016/0003-9861(62)90412-5. PMID 14482339.
- ^ Cleaves II, Henderson (2010). "The origin of the biologically coded amino acids". Journal of Theoretical Biology. 263 (4): 490-498. Bibcode:2010JThBi.263..490C. doi:10.1016/j.jtbi.2009.12.014. PMID 20034500.
- ^ Breslow, R. (1959). "On the Mechanism of the Formose Reaction". Tetrahedron Letters. 1 (21): 22-26. doi:10.1016/S0040-4039(01)99487-0.
- ^ Oró, Joan (16 Eylül 1961). "Mechanism of Synthesis of Adenine from Hydrogen Cyanide under Possible Primitive Earth Conditions". Nature. 191 (4794): 1193-1194. Bibcode:1961Natur.191.1193O. doi:10.1038/1911193a0. PMID 13731264.
- ^ a b Saladino, Raffaele; Crestini, Claudia; Pino, Samanta; Costanzo, Giovanna; Di Mauro, Ernesto (March 2012). "Formamide and the origin of life" (PDF). Physics of Life Reviews. 9 (1): 84-104. Bibcode:2012PhLRv...9...84S. doi:10.1016/j.plrev.2011.12.002. hdl:2108/85168. PMID 22196896. 27 Ocak 2023 tarihinde kaynağından arşivlendi (PDF). Erişim tarihi: 2 Nisan 2023.
- ^ a b Saladino, Raffaele; Botta, Giorgia; Pino, Samanta; Costanzo, Giovanna; Di Mauro, Ernesto (July 2012). "From the one-carbon amide formamide to RNA all the steps are prebiotically possible". Biochimie. 94 (7): 1451-1456. doi:10.1016/j.biochi.2012.02.018. hdl:11573/515604. PMID 22738728.
- ^ Marlaire, Ruth, (Ed.) (3 Mart 2015). "NASA Ames Reproduces the Building Blocks of Life in Laboratory". Ames Research Center. NASA. 5 Mart 2015 tarihinde kaynağından arşivlendi. Erişim tarihi: 5 Mart 2015.
- ^ Ferus, Martin; Nesvorný, David; Šponer, Jiří; Kubelík, Petr; Michalčíková, Regina; Shestivská, Violetta; Šponer, Judit E.; Civiš, Svatopluk (2015). "High-energy chemistry of formamide: A unified mechanism of nucleobase formation". PNAS. 112 (3): 657-662. Bibcode:2015PNAS..112..657F. doi:10.1073/pnas.1412072111. PMC 4311869 $2. PMID 25489115.
- ^ Basile, Brenda; Lazcano, Antonio; Oró, Joan (1984). "Prebiotic syntheses of purines and pyrimidines". Advances in Space Research. 4 (12): 125-131. Bibcode:1984AdSpR...4l.125B. doi:10.1016/0273-1177(84)90554-4. PMID 11537766.
- ^ Orgel, Leslie E. (August 2004). "Prebiotic Adenine Revisited: Eutectics and Photochemistry". Origins of Life and Evolution of Biospheres. 34 (4): 361-369. Bibcode:2004OLEB...34..361O. doi:10.1023/B:ORIG.0000029882.52156.c2. PMID 15279171.
- ^ Robertson, Michael P.; Miller, Stanley L. (29 Haziran 1995). "An efficient prebiotic synthesis of cytosine and uracil". Nature. 375 (6534): 772-774. Bibcode:1995Natur.375..772R. doi:10.1038/375772a0. PMID 7596408.
- ^ Fox, Douglas (1 Şubat 2008). "Did Life Evolve in Ice?". Discover. 30 Haziran 2008 tarihinde kaynağından arşivlendi. Erişim tarihi: 3 Temmuz 2008.
- ^ Levy, Matthew; Miller, Stanley L.; Brinton, Karen; Bada, Jeffrey L. (June 2000). "Prebiotic Synthesis of Adenine and Amino Acids Under Europa-like Conditions". Icarus. 145 (2): 609-613. Bibcode:2000Icar..145..609L. doi:10.1006/icar.2000.6365. PMID 11543508.
- ^ Menor-Salván, César; Ruiz-Bermejo, Marta; Guzmán, Marcelo I.; Osuna-Esteban, Susana; Veintemillas-Verdaguer, Sabino (20 Nisan 2009). "Synthesis of Pyrimidines and Triazines in Ice: Implications for the Prebiotic Chemistry of Nucleobases". Chemistry: A European Journal. 15 (17): 4411-4418. doi:10.1002/chem.200802656. PMID 19288488.
- ^ Roy, Debjani; Najafian, Katayoun; von Ragué Schleyer, Paul (30 Ekim 2007). "Chemical evolution: The mechanism of the formation of adenine under prebiotic conditions". PNAS. 104 (44): 17272-17277. Bibcode:2007PNAS..10417272R. doi:10.1073/pnas.0708434104. PMC 2077245 $2. PMID 17951429.
- ^ Lancet, Doron (30 Aralık 2014). "Systems Prebiology-Studies of the origin of Life". The Lancet Lab. Rehovot, Israel: Department of Molecular Genetics; Weizmann Institute of Science. 26 Haziran 2015 tarihinde kaynağından arşivlendi. Erişim tarihi: 26 Haziran 2015.
- ^ Segré, Daniel; Ben-Eli, Dafna; Deamer, David W.; Lancet, Doron (February 2001). "The Lipid World" (PDF). Origins of Life and Evolution of Biospheres. 31 (1–2): 119-145. Bibcode:2001OLEB...31..119S. doi:10.1023/A:1006746807104. PMID 11296516. 26 Haziran 2015 tarihinde kaynağından arşivlendi (PDF).
- ^ a b c Chen, Irene A.; Walde, Peter (July 2010). "From Self-Assembled Vesicles to Protocells". Cold Spring Harbor Perspectives in Biology. 2 (7): a002170. doi:10.1101/cshperspect.a002170. PMC 2890201 $2. PMID 20519344.
- ^ Eigen, Manfred; Schuster, Peter (November 1977). "The Hypercycle. A Principle of Natural Self-Organization. Part A: Emergence of the Hypercycle" (PDF). Naturwissenschaften. 64 (11): 541-65. Bibcode:1977NW.....64..541E. doi:10.1007/bf00450633. PMID 593400. 3 Mart 2016 tarihinde kaynağından (PDF) arşivlendi.
- ^ Markovitch, Omer; Lancet, Doron (Yaz 2012). "Excess Mutual Catalysis Is Required for Effective Evolvability". Artificial Life. 18 (3): 243-266. doi:10.1162/artl_a_00064. PMID 22662913.
- ^ Tessera, Marc (2011). "Origin of Evolution versus Origin of Life: A Shift of Paradigm". International Journal of Molecular Sciences. 12 (6): 3445-3458. doi:10.3390/ijms12063445. PMC 3131571 $2. PMID 21747687. Special Issue: "Origin of Life 2011"
- ^ Onsager, Lars (1931). "Reciprocal Relations in Irreversible Processes I and II". Physical Review. 37 (4): 405. Bibcode:1931PhRv...37..405O. doi:10.1103/PhysRev.37.405.
- ^ Onsager, Lars (1931). "Reciprocal Relations in Irreversible Processes I and II". Physical Review (38): 2265. doi:10.1103/PhysRev.38.2265.
- ^ Prigogine, Ilya (1967). An Introduction to the Thermodynamics of Irreversible Processes. New York: Wiley.
- ^ "Exploring Life's Origins: Protocells". Exploring Life's Origins: A Virtual Exhibit. Arlington County, Virginia: National Science Foundation. 28 Şubat 2014 tarihinde kaynağından arşivlendi. Erişim tarihi: 18 Mart 2014.
- ^ a b c Chen, Irene A. (8 Aralık 2006). "The Emergence of Cells During the Origin of Life". Science. 314 (5805): 1558-1559. doi:10.1126/science.1137541. PMID 17158315.
- ^ Zimmer, Carl (26 Haziran 2004). "What Came Before DNA?". Discover. 19 Mart 2014 tarihinde kaynağından arşivlendi.
- ^ Shapiro, Robert (June 2007). "A Simpler Origin for Life". Scientific American. 296 (6): 46-53. Bibcode:2007SciAm.296f..46S. doi:10.1038/scientificamerican0607-46. PMID 17663224. 14 Haziran 2015 tarihinde kaynağından arşivlendi.
- ^ Chang 2007
- ^ a b c d Lane, Nick (2015). The Vital Question: Why Is Life The Way It Is?. Profile Books. ss. 129-140. ISBN 978-1781250365.
- ^ Sharov, Alexei A.; Gordon, Richard (2018). "Life Before Earth". Habitability of the Universe Before Earth: Life Before Earth. Astrobiology Exploring Life on Earth and Beyond. Academic Press. ss. 265-296. doi:10.1016/B978-0-12-811940-2.00011-3. ISBN 9780128119402.
- ^ Ladyman, J.; Lambert, J.; Weisner, K. B. (2013). "What is a Complex System?". European Journal of the Philosophy of Science. 3: 33-67. doi:10.1007/s13194-012-0056-8.
- ^ Esposito, M.; Lindenberg, Katja; Van den Broeck, C. (2010). "Entropy production as correlation between system and reservoir". New Journal of Physics. 12 (1): 013013. arXiv:0908.1125 $2. Bibcode:2010NJPh...12a3013E. doi:10.1088/1367-2630/12/1/013013.
- ^ Bar-Nun, A.; Bar-Nun, N.; Bauer, S. H.; Sagan, Carl (24 Nisan 1970). "Shock Synthesis of Amino Acids in Simulated Primitive Environments". Science. 168 (3930): 470-473. Bibcode:1970Sci...168..470B. doi:10.1126/science.168.3930.470. PMID 5436082.
- ^ Anbar, Michael (27 Eylül 1968). "Cavitation during Impact of Liquid Water on Water: Geochemical Implications". Science. 161 (3848): 1343-1344. Bibcode:1968Sci...161.1343A. doi:10.1126/science.161.3848.1343. PMID 17831346.
- ^ Dharmarathne, Leena; Grieser, Franz (7 Ocak 2016). "Formation of Amino Acids on the Sonolysis of Aqueous Solutions Containing Acetic Acid, Methane, or Carbon Dioxide, in the Presence of Nitrogen Gas". The Journal of Physical Chemistry A. 120 (2): 191-199. Bibcode:2016JPCA..120..191D. doi:10.1021/acs.jpca.5b11858. PMID 26695890.
- ^ Patehebieke, Yeersen; Zhao, Ze-Run; Wang, Su; Xu, Hao-Xing; Chen, Qian-Qian; Wang, Xiao (2021). "Cavitation as a plausible driving force for the prebiotic formation of N9 purine nucleosides". Cell Reports Physical Science. 2 (3): 100375. Bibcode:2021CRPS....200375P. doi:10.1016/j.xcrp.2021.100375.
- ^ Kalson, Natan-Haim; Furman, David; Zeiri, Yehuda (11 Eylül 2017). "Cavitation-Induced Synthesis of Biogenic Molecules on Primordial Earth". ACS Central Science. 3 (9): 1041-1049. doi:10.1021/acscentsci.7b00325. PMC 5620973 $2. PMID 28979946.
- ^ Muller, Anthonie W. J. (1995). "Were the first organisms heat engines? A new model for biogenesis and the early evolution of biological energy conversion". Progress in Biophysics and Molecular Biology. 63 (2): 193-231. doi:10.1016/0079-6107(95)00004-7. PMID 7542789.
- ^ Muller, Anthonie W. J.; Schulze-Makuch, Dirk (2006). "Thermal energy and the origin of life". Origins of Life and Evolution of Biospheres. 36 (2): 77-189. Bibcode:2006OLEB...36..177M. doi:10.1007/s11084-005-9003-4. PMID 16642267.
- ^ Junge, Wolfgang; Nelson, Nathan (2 Haziran 2015). "ATP Synthase". Annual Review of Biochemistry. 84 (1): 631-657. doi:10.1146/annurev-biochem-060614-034124. PMID 25839341.
- ^ a b c Damer, Bruce; Deamer, David (1 Nisan 2020). "The Hot Spring Hypothesis for an Origin of Life". Astrobiology. 20 (4): 429-452. Bibcode:2020AsBio..20..429D. doi:10.1089/ast.2019.2045. ISSN 1531-1074. PMC 7133448 $2. PMID 31841362.
- ^ Benner, S. A.; Bell, E. A.; Biondi, E.; Brasser, R.; Carell, T.; Kim, H.-J.; Mojzsis, S. J.; Omran, A.; Pasek, M. A.; Trail, D. (2020). "When Did Life Likely Emerge on Earth in an RNA-First Process?". ChemSystemsChem. 2 (2). doi:10.1002/syst.201900035.
- ^ Copley, Shelley D.; Smith, Eric; Morowitz, Harold J. (December 2007). "The origin of the RNA world: Co-evolution of genes and metabolism" (PDF). Bioorganic Chemistry. 35 (6): 430-443. doi:10.1016/j.bioorg.2007.08.001. PMID 17897696. 5 Eylül 2013 tarihinde kaynağından arşivlendi (PDF). Erişim tarihi: 8 Haziran 2015.
The proposal that life on Earth arose from an RNA world is widely accepted.
- ^ a b c Robertson, Michael P.; Joyce, Gerald F. (May 2012). "The origins of the RNA world". Cold Spring Harbor Perspectives in Biology. 4 (5): a003608. doi:10.1101/cshperspect.a003608. PMC 3331698 $2. PMID 20739415.
- ^ a b c Cech, Thomas R. (July 2012). "The RNA Worlds in Context". Cold Spring Harbor Perspectives in Biology. 4 (7): a006742. doi:10.1101/cshperspect.a006742. PMC 3385955 $2. PMID 21441585.
- ^ Pearce, Ben K. D.; Pudritz, Ralph E.; Semenov, Dmitry A.; Henning, Thomas K. (24 Ekim 2017). "Origin of the RNA world: The fate of nucleobases in warm little ponds". PNAS. 114 (43): 11327-11332. arXiv:1710.00434 $2. Bibcode:2017PNAS..11411327P. doi:10.1073/pnas.1710339114. PMC 5664528 $2. PMID 28973920.
- ^ Yarus, Michael (April 2011). "Getting Past the RNA World: The Initial Darwinian Ancestor". Cold Spring Harbor Perspectives in Biology. 3 (4): a003590. doi:10.1101/cshperspect.a003590. PMC 3062219 $2. PMID 20719875.
- ^ Voet & Voet 2004, s. 29
- ^ Fox, George.E. (9 Haziran 2010). "Origin and evolution of the ribosome". Cold Spring Harbor Perspectives in Biology. 2 (9(a003483)): a003483. doi:10.1101/cshperspect.a003483. PMC 2926754 $2. PMID 20534711.
- ^ Neveu, Marc; Kim, Hyo-Joong; Benner, Steven A. (22 Nisan 2013). "The 'Strong' RNA World Hypothesis: Fifty Years Old". Astrobiology. 13 (4): 391-403. Bibcode:2013AsBio..13..391N. doi:10.1089/ast.2012.0868. PMID 23551238.
- ^ Gilbert, Walter (20 Şubat 1986). "Origin of life: The RNA world". Nature. 319 (6055): 618. Bibcode:1986Natur.319..618G. doi:10.1038/319618a0.
- ^ Orgel, Leslie E. (October 1994). "The origin of life on Earth". Scientific American. 271 (4): 76-83. Bibcode:1994SciAm.271d..76O. doi:10.1038/scientificamerican1094-76. PMID 7524147.
- ^ Lincoln, Tracey A.; Joyce, Gerald F. (27 Şubat 2009). "Self-Sustained Replication of an RNA Enzyme". Science. 323 (5918): 1229-1232. Bibcode:2009Sci...323.1229L. doi:10.1126/science.1167856. PMC 2652413 $2. PMID 19131595.
- ^ Joyce, Gerald F. (2009). "Evolution in an RNA world". Cold Spring Harbor Perspectives in Biology. 74 (Evolution: The Molecular Landscape): 17-23. doi:10.1101/sqb.2009.74.004. PMC 2891321 $2. PMID 19667013.
- ^ Szostak, Jack W. (5 Şubat 2015). "The Origins of Function in Biological Nucleic Acids, Proteins, and Membranes". Chevy Chase, Maryland: Howard Hughes Medical Institute. 14 Temmuz 2015 tarihinde kaynağından arşivlendi. Erişim tarihi: 16 Haziran 2015.
- ^ Bernstein, Harris; Byerly, Henry C.; Hopf, Frederick A.; Michod, Richard A.; Vemulapalli, G. Krishna (June 1983). "The Darwinian Dynamic". The Quarterly Review of Biology. 58 (2): 185-207. doi:10.1086/413216. JSTOR 2828805.
- ^ Michod 1999
- ^ Palasek, Stan (23 Mayıs 2013). "Primordial RNA Replication and Applications in PCR Technology". arXiv:1305.5581v1 $2.
- ^ Vlassov, Alexander V.; Kazakov, Sergei A.; Johnston, Brian H.; Landweber, Laura F. (August 2005). "The RNA World on Ice: A New Scenario for the Emergence of RNA Information". Journal of Molecular Evolution. 61 (2): 264-273. Bibcode:2005JMolE..61..264V. doi:10.1007/s00239-004-0362-7. PMID 16044244.
- ^ Nussinov, Mark D.; Otroshchenko, Vladimir A.; Santoli, Salvatore (1997). "The emergence of the non-cellular phase of life on the fine-grained clayish particles of the early Earth's regolith". BioSystems. 42 (2–3): 111-118. doi:10.1016/S0303-2647(96)01699-1. PMID 9184757.
- ^ Kühnlein, Alexandra; Lanzmich, Simon A.; Brun, Dieter (2 Mart 2021). "tRNA sequences can assemble into a replicator". eLife. 10. doi:10.7554/eLife.63431. PMC 7924937 $2. PMID 33648631.
- ^ Noller, Harry F. (April 2012). "Evolution of protein synthesis from an RNA world". Cold Spring Harbor Perspectives in Biology. 4 (4): a003681. doi:10.1101/cshperspect.a003681. PMC 3312679 $2. PMID 20610545.
- ^ Koonin, Eugene V. (31 Mayıs 2007). "The cosmological model of eternal inflation and the transition from chance to biological evolution in the history of life". Biology Direct. 2: 15. doi:10.1186/1745-6150-2-15. PMC 1892545 $2. PMID 17540027.
- ^ Boone, David R.; Castenholz, Richard W.; Garrity, George M., (Ed.) (2001). The Archaea and the Deeply Branching and Phototrophic Bacteria. Bergey's Manual of Systematic Bacteriology. Springer. ISBN 978-0-387-21609-6. 25 Aralık 2014 tarihinde kaynağından arşivlendi.[sayfa belirt]
- ^ Woese, C. R.; Fox, G. E. (1977). "Phylogenetic structure of the prokaryotic domain: the primary kingdoms". PNAS. 7 (11): 5088-5090. Bibcode:1977PNAS...74.5088W. doi:10.1073/pnas.74.11.5088. PMC 432104 $2. PMID 270744.
- ^ Valas, R. E.; Bourne, P. E. (2011). "The origin of a derived superkingdom: how a gram-positive bacterium crossed the desert to become an archaeon". Biology Direct. 6: 16. doi:10.1186/1745-6150-6-16. PMC 3056875 $2. PMID 21356104.
- ^ Cavalier-Smith, Thomas (2006). "Rooting the tree of life by transition analyses". Biology Direct. 1: 19. doi:10.1186/1745-6150-1-19. PMC 1586193 $2. PMID 16834776.
- ^ "Early life liked it hot". Nature. 535 (7613): 468. 2016. doi:10.1038/535468b.
- ^ Gogarten, Johann Peter; Deamer, David (25 Kasım 2016). "Is LUCA a thermophilic progenote?". Nature Microbiology (İngilizce). 1 (12): 16229. doi:10.1038/nmicrobiol.2016.229. ISSN 2058-5276. PMID 27886195. 3 Nisan 2020 tarihinde kaynağından arşivlendi. Erişim tarihi: 2 Nisan 2023.
- ^ Catchpole, Ryan; Forterre, Patrick (2019). "The evolution of Reverse Gyrase suggests a non-hyperthermophilic Last Universal Common Ancestor". Molecular Biology and Evolution. 36 (12): 2737-2747. doi:10.1093/molbev/msz180. PMC 6878951 $2. PMID 31504731. 27 Ocak 2023 tarihinde kaynağından arşivlendi. Erişim tarihi: 2 Nisan 2023.
- ^ Berkemer, Sarah J.; McGlynn, Shawn E (8 Ağustos 2020). "A New Analysis of Archaea–Bacteria Domain Separation: Variable Phylogenetic Distance and the Tempo of Early Evolution". Molecular Biology and Evolution. 37 (8): 2332-2340. doi:10.1093/molbev/msaa089. PMC 7403611 $2. PMID 32316034. 27 Ocak 2023 tarihinde kaynağından arşivlendi. Erişim tarihi: 2 Nisan 2023.
- ^ Hoffmann, Geoffrey W. (25 Haziran 1974). "On the origin of the genetic code and the stability of the translation apparatus". Journal of Molecular Biology. 86 (2): 349-362. doi:10.1016/0022-2836(74)90024-2. PMID 4414916.
- ^ Orgel, Leslie E. (April 1963). "The Maintenance of the Accuracy of Protein Synthesis and its Relevance to Ageing". PNAS. 49 (4): 517-521. Bibcode:1963PNAS...49..517O. doi:10.1073/pnas.49.4.517. PMC 299893 $2. PMID 13940312.
- ^ Hoffmann, Geoffrey W. (October 1975). "The Stochastic Theory of the Origin of the Genetic Code". Annual Review of Physical Chemistry. 26: 123-144. Bibcode:1975ARPC...26..123H. doi:10.1146/annurev.pc.26.100175.001011.
- ^ Brasier, M. D. (2012). Secret Chambers: The Inside Story of Cells and Complex Life. Oxford University Press. s. 298.
- ^ Ward, Peter & Kirschvink, Joe, op cit, p. 42
- ^ a b Colín-García, M.; Heredia, A.; Cordero, G.; Camprubí, A.; Negrón-Mendoza, A.; Ortega-Gutiérrez, F.; Berald, H.; Ramos-Bernal, S. (2016). "Hydrothermal vents and prebiotic chemistry: a review". Boletín de la Sociedad Geológica Mexicana. 68 (3): 599-620. doi:10.18268/BSGM2016v68n3a13. 18 Ağustos 2017 tarihinde kaynağından arşivlendi.
- ^ Schirber, Michael (24 Haziran 2014). "Hydrothermal Vents Could Explain Chemical Precursors to Life". NASA Astrobiology: Life in the Universe. NASA. 29 Kasım 2014 tarihinde kaynağından arşivlendi. Erişim tarihi: 19 Haziran 2015.
- ^ a b Martin, William; Russell, Michael J. (29 Ocak 2003). "On the origins of cells: a hypothesis for the evolutionary transitions from abiotic geochemistry to chemoautotrophic prokaryotes, and from prokaryotes to nucleated cells". Philosophical Transactions of the Royal Society B. 358 (1429): 59-83; discussion 83-85. doi:10.1098/rstb.2002.1183. PMC 1693102 $2. PMID 12594918.
- ^ a b Lane 2009
- ^ Usher, Oli (27 Nisan 2015). "Chemistry of seabed's hot vents could explain emergence of life" (Basın açıklaması). University College London. 20 Haziran 2015 tarihinde kaynağından arşivlendi. Erişim tarihi: 19 Haziran 2015.
- ^ Roldan, Alberto; Hollingsworth, Nathan; Roffey, Anna; Islam, Husn-Ubayda; Goodall, Josephine B. M.; Catlow, C. Richard A.; Darr, Jawwad A.; Bras, Wim; Sankar, Gopinathan; Holt, Katherine B.; Hogarth, Graeme; de Leeuw, Nora Henriette (May 2015). "Bio-inspired CO2 conversion by iron sulfide catalysts under sustainable conditions". Chemical Communications. 51 (35): 7501-7504. doi:10.1039/C5CC02078F. PMID 25835242. 20 Haziran 2015 tarihinde kaynağından arşivlendi. Erişim tarihi: 19 Haziran 2015.
- ^ Baross, J. A.; Hoffman, S. E. (1985). "Submarine hydrothermal vents and associated gradient environments as sites for the origin and evolution of life". Origins of Life and Evolution of Biospheres. 15 (4): 327-345. Bibcode:1985OrLi...15..327B. doi:10.1007/bf01808177.
- ^ Russell, M. J.; Hall, A. J. (1997). "The emergence of life from iron monosulphide bubbles at a submarine hydrothermal redox and pH front". Journal of the Geological Society. 154 (3): 377-402. Bibcode:1997JGSoc.154..377R. doi:10.1144/gsjgs.154.3.0377. PMID 11541234.
- ^ Amend, J. P.; LaRowe, D. E.; McCollom, T. M.; Shock, E. L. (2013). "The energetics of organic synthesis inside and outside the cell". Philosophical Transactions of the Royal Society B. 368 (1622): 20120255. doi:10.1098/rstb.2012.0255. PMC 3685458 $2. PMID 23754809.
- ^ Shock, E. L.; Boyd, E. S. (2015). "Geomicrobiology and microbial geochemistry:principles of geobiochemistry". Elements. 11: 389-394. doi:10.2113/gselements.11.6.395.
- ^ Martin, W.; Russell, M. J. (2007). "On the origin of biochemistry at an alkaline hydrothermal vent". Philosophical Transactions of the Royal Society B. 362 (1486): 1887-1925. doi:10.1098/rstb.2006.1881. PMC 2442388 $2. PMID 17255002.
- ^ Weiss, Madeline C.; Sousa, Filipa L.; Mrnjavac, Natalia; Neukirchen, Sinje; Roettger, Mayo; Nelson-Sathi, Shijulal; Martin, William F. (25 Temmuz 2016). "The physiology and habitat of the last universal common ancestor" (PDF). Nature Microbiology. 1 (9): 16116. doi:10.1038/nmicrobiol.2016.116. PMID 27562259. 18 Nisan 2022 tarihinde kaynağından arşivlendi (PDF). Erişim tarihi: 2 Nisan 2023.
- ^ Lane, Nick; Martin, William F. (21 Aralık 2012). "The Origin of Membrane Bioenergetics". Cell (İngilizce). 151 (7): 1406-1416. doi:10.1016/j.cell.2012.11.050. ISSN 0092-8674. PMID 23260134. 6 Aralık 2022 tarihinde kaynağından arşivlendi. Erişim tarihi: 2 Nisan 2023.
- ^ Baaske, Philipp; Weinert, Franz M.; Duhr, Stefan; Lemke, Kono H.; Russell, Michael J.; Braun, Dieter (29 Mayıs 2007). "Extreme accumulation of nucleotides in simulated hydrothermal pore systems". Proceedings of the National Academy of Sciences (İngilizce). 104 (22): 9346-9351. doi:10.1073/pnas.0609592104. ISSN 0027-8424. PMC 1890497 $2. PMID 17494767.
- ^ a b c Mulkidjanian, Armen Y.; Bychkov, Andrew Yu.; Dibrova, Daria V.; Galperin, Michael Y.; Koonin, Eugene V. (3 Nisan 2012). "Origin of first cells at terrestrial, anoxic geothermal fields". Proceedings of the National Academy of Sciences. 109 (14): E821-30. Bibcode:2012PNAS..109E.821M. doi:10.1073/pnas.1117774109. PMC 3325685 $2. PMID 22331915.
- ^ Chandru, Kuhan; Guttenberg, Nicholas; Giri, Chaitanya; Hongo, Yayoi; Butch, Christopher; Mamajanov, Irena; Cleaves, H. James (31 Mayıs 2018). "Simple prebiotic synthesis of high diversity dynamic combinatorial polyester libraries". Communications Chemistry. 1 (1). doi:10.1038/s42004-018-0031-1.
- ^ Forsythe, Jay G.; Yu, Sheng-Sheng; Mamajanov, Irena; Grover, Martha A; Krishnamurthy, Ramanarayanan; Fernández, Facundo M.; Hud, Nicholas V. (17 Ağustos 2015). "Ester-Mediated Amide Bond Formation Driven by Wet–Dry Cycles: A Possible Path to Polypeptides on the Prebiotic Earth". Angewandte Chemie International Edition in English. 54 (34): 9871-9875. doi:10.1002/anie.201503792. PMC 4678426 $2. PMID 26201989.
- ^ Patel, Bhavesh H.; Percivalle, Claudia; Ritson, Dougal J.; Duffy, Colm. D.; Sutherland, John D. (16 Mart 2015). "Common origins of RNA, protein and lipid precursors in a cyanosulfidic protometabolism". Nature Chemistry. 7 (4): 301-307. Bibcode:2015NatCh...7..301P. doi:10.1038/nchem.2202. ISSN 1755-4330. PMC 4568310 $2. PMID 25803468.
- ^ Deamer, David (10 Şubat 2021). "Where Did Life Begin? Testing Ideas in Prebiotic Analogue Conditions". Life. 11 (2): 134. Bibcode:2021Life...11..134D. doi:10.3390/life11020134. PMC 7916457 $2. PMID 33578711.
- ^ Cairns-Smith, Graham (2 Eylül 1982). Genetic Takeover and the Mineral Origins of Life. Cambridge: Cambridge University Press. ISBN 0-521-23312-7. OCLC 7875600.
- ^ Dawkins 1996, ss. 148–161
- ^ Huang, Wenhua; Ferris, James P. (12 Temmuz 2006). "One-Step, Regioselective Synthesis of up to 50-mers of RNA Oligomers by Montmorillonite Catalysis". Journal of the American Chemical Society. 128 (27): 8914-8919. doi:10.1021/ja061782k. PMID 16819887.