
 French
French Deutsch
DeutschЛабиринтулы — Википедия
| Лабиринтулы | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
 Вегетативная клетка и эктоплазматическая сеть Aplanochytrium sp. | ||||||||||||
| Научная классификация | ||||||||||||
| Международное научное название | ||||||||||||
| Labyrinthulea Olive, 1975 | ||||||||||||
| Синонимы | ||||||||||||
| ||||||||||||
| Дочерние таксоны | ||||||||||||
| См. раздел Систематика | ||||||||||||
| ||||||||||||
Лабири́нтулы (лат. Labyrinthulea, МКЗН), или лабиринтуломице́ты (лат. Labyrinthulomycetes, МКН)[1][2][3] — класс протистов, относящихся к надтипу Heterokonta, или Stramenopiles. Включает около 40 видов — преимущественно морских гетеротрофных грибоподобных организмов[2].
В русскоязычной литературе укоренилась традиция[4] именовать термином «лабиринтулы» как представителей класса Labyrinthulea в целом, так и представителей входящего в него рода Labyrinthula (в данной статье упомянутый термин употребляется в широком значении, а там, где речь заходит именно о роде Labyrinthula, это указано явно).
Лабиринтулы распространены в океанах повсеместно[5]. Вегетативная стадия у большинства видов представлена плазмодием (сетчатым или ризоидоподобным), состоящим из веретеновидных или овальных клеток, которые погружены в эктоплазматическую сеть, волокна которой покрыты мембраной[1][6].
С точки зрения экологии лабиринтулы — сапротрофные редуценты, осуществляющие круговорот органики в морских и прибрежных экосистемах, разлагая детрит внеклеточными ферментами. Некоторые лабиринтулы вызывают заболевания животных и растений. Другие представители служат источниками ценных химических веществ — например, длинноцепочечных омега-3-полиненасыщенных жирных кислот, которые используются в качестве пищевых добавок, а потому имеют важное промышленное значение[7].
История изучения
[править | править код]
Изучение лабиринтул началось в 1867 году, когда российский ботаник Л. С. Ценковский описал[8] два вида, вошедших в выделенный им род лабиринтулы (Labyrinthula) — первый из родов, в настоящее время относимых к данному классу. Обнаруженные им лабиринтулы произрастали на морских водорослях, найденных в Чёрном море неподалёку от Одессы. Положение данного рода (выделенного в 1868 немецким естествоиспытателем Э. Геккелем в отдельное семейство) в системе органического мира долгое время оставалось неясным — их относили и к миксомицетам, и к грибам, и к простейшим[9].
В 1936 году американский миколог Ф. К. Спэрроу, исследуя морские грибы, встречавшиеся в окрестностях местечка Вудс-Хол[англ.] (штат Массачусетс), описал[10] новый род траустохитриум (Thraustochytrium) с видом Thraustochytrium proliferum, а в 1943 году выделил его в отдельное семейство, отнесённое к оомицетам (его обычно включали, хотя и с оговорками, в состав порядка сапролегниальные[11]). Позднее были описаны и другие роды данного семейства[1][12].
Оба семейства первоначально рассматривались как не связанные друг с другом. Однако проведённые в 1950—1960-х годах ультраструктурные исследования показали структурную близость обеих групп (наличие внеклеточной плазматической сети и ботросом, сходство в строении клеточных стенок, зооспор и их жгутикового аппарата), что позволило американскому микологу Л. Оливу объединить в 1975 году оба семейства в единый таксон — класс Labyrinthulea (Лабиринтулы), содержащий один отряд и два семейства: Labyrinthulidae (лабиринтуловые; иногда в русскоязычной литературе данную группу протистов именуют также сетчатые слизевики[13]) и Thraustochytridae (траустохитриевые)[1][14]. В 1982 году Олайв счёл необходимым повысить ранг этих двух подразделений лабиринтул до ранга отряда[15]. Формального латинского диагноза для данного класса он, однако, не дал; это упущение было исправлено в 1986 году английским биологом Т. Кавалье-Смитом, который со всей определённостью отнёс лабиринтул к группе гетероконтов[16].
В 2012 году О. Р. Андерсон и Т. Кавалье-Смит пересмотрели структуру класса Labyrinthulea, включив в его состав роды Diplophrys, Sorodiplophrys, Amphifila (ранее относимые к протистам из группы Cercozoa) и подразделив класс на 2 отряда и 8 семейств (см. далее)[17].
Морфология
[править | править код]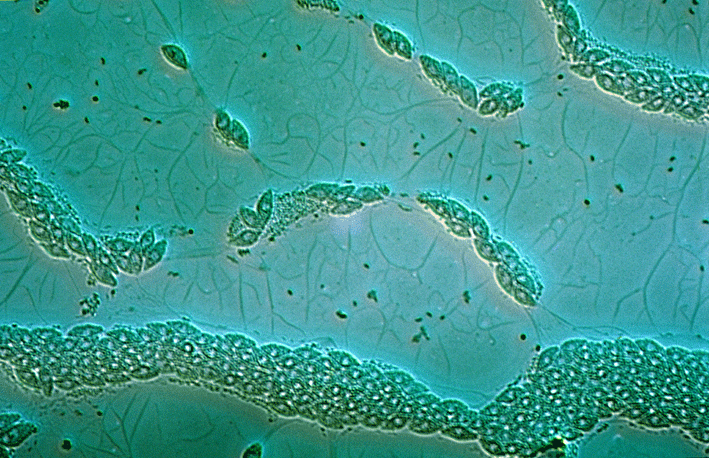{kind=link}
{kind=link}
{kind=link}
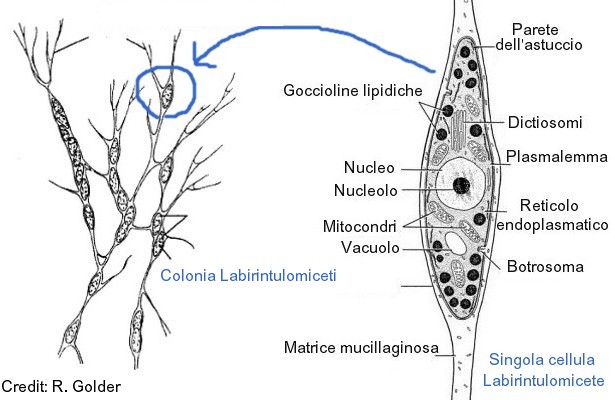
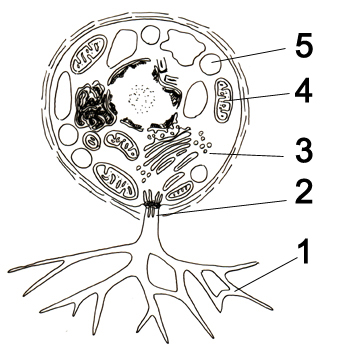
Своеобразие морфологии лабиринтул наиболее чётко можно проиллюстрировать на примере рода лабиринтулы (Labyrinthula). На вегетативной стадии (стадии трофонта) представители данного рода существуют в виде блуждающих сетчатых плазмодиев: веретеновидные бесцветные или желтоватые тельца, содержащие ядро и имеющие постоянную или изменчивую форму («клетки»), скользят внутри составляющих сеть слизистых трубок, похожих на псевдоподии. Совокупность таких слизистых тяжей и представляет собой у представители рода Labyrinthula эктоплазматическую сеть. Слизь, заполняющая трубки, состоит из полисахаридов, и «клетки» могут свободно скользить по ней, а покровом эктоплазматической сети выступает плазмалемма. Каждая веретеновидная «клетка» окружена уплощённой оболочкой, состоящей из двух параллельных мембран с небольшим просветом между ними, в котором располагаются чешуйки. Чешуйки круглые, образуются в диктиосомах[18]. В одной или нескольких точках две мембраны соединяются, образуя пору — ботросому (сагеногенетосома, сагеноген), так что оболочка не является сплошной. Ультраструктура ботросомы очень сложна; через ботросомы цитоплазмы «клеток» сообщаются с цитоплазмой сети. Наличие оболочки из двух мембран и ботросомы — уникальный (апоморфный) признак всего класса лабиринтул (хотя у отдельных представителей он вторично утрачен). «Клетки» по сути не являются истинными клетками, поскольку каждый содержащий ядро участок лишь частично отграничен от обобществлённой сети цитоплазматических тяжей[4][13].
Если у Labyrinthulidae плазмодий является сетчатым и его слизистые тяжи окружают каждую «клетку», то у Aplanochytridae и большинства представителей Thraustochytrida плазмодий образует древовидную структуру (из спорангия, окружённого плотной стенкой, исходят ветвящиеся слизистые тяжи), напоминая ризомицелий хитридиевых грибов[19]. Андерсон и Кавалье-Смит выделяют у лабиринтул три основных морфологических типа. Первый тип, характерный для большинства траустохитрид, предполагает наличие неподвижных вегетативных клеток с одной ботросомой, из которой исходят тонкие ветвящиеся филоподии[англ.] (служащие не для передвижения, а, вероятно, для увеличения площади поверхности клетки с созданием лучших условий поглощения растворённых в окружающей среде органических молекул), и двужгутиковых зооспор. Второй тип, характерный для лабиринтулид, отличается наличием сетчатого плазмодия с неподвижными (у Aplanochytriidae) или скользящими в нём (у Labyrinthulidae) вегетативными клетками, а также зооспор. Третий тип, который характерен для Diplophrys и сходных с ним родов, предполагает наличие двух полярных пучков филоподий и отсутствие эктоплазматической сети, ботросом и зооспор[17].
Эктоплазматическая сеть не только обеспечивает прикрепление к субстрату[18] и движение «клеток», но и направляет их движение в сторону источника питания, обеспечивает перемещение веществ между «клетками» и окружающей средой и совместное существование «клеток», защищает «клетки» от неблагоприятных внешних факторов (например, высыхания) и позволяет плазмодиям выползать из воды на сушу[13]. Механизм движения «клеток» неизвестен (вероятно, при движении задействованы расположенные в сети актиновые филаменты[20] и мукополисахариды). Сокращение актиновых филаментов в слизистых тяжах зависит от концентрации ионов кальция. По-видимому, сила, создаваемая движением актинового филамента вдоль слизистого слоя, лежащего над «клетками», заставляет «клетки» двигаться вперёд со скоростью несколько микрометров в секунду. В состоянии покоя движения хаотичны, однако при появлении питательных веществ (аттрактантов) становятся чётко направленными в их сторону[21].
Описанная выше эктоплазматическая сеть характерна не для всех представителей класса лабиринтул; в частности, она отсутствует у Althornia[18]. Некоторые лабиринтулы имеют талломы, внешне напоминающие талломы хитридиомицетов, однако это сходство лишь поверхностное[6].
Клеточная биология
[править | править код]
Стенка спорангия[21] лабиринтул содержит белки и простейшие сахара (как правило, галактозу, однако у некоторых видов преобладают фукоза, рамноза или ксилоза). Считается, что использование фукозы нерационально для клеток, поэтому наличие этого сахара у лабиринтул может говорить в пользу их эволюционной древности. Лабиринтулы не способны синтезировать аминокислоту лизин. Основное запасное вещество — гликоген[22]. Многие представители класса накапливают в клетках большие количества омега-3-полиненасыщенных жирных кислот — таких, как эйкозапентаеновая кислота, докозагексаеновая кислота и докозапентаеновая кислота[англ.][5]. Кристы митохондрий трубчатые[18].
Несмотря на то, что лабиринтулы относятся к страменопилам, их питание во многом подобно питанию грибов. Лабиринтулы питаются осмотрофно, выпуская ризоиды над детритом, и выделяют наружу ферменты, расщепляющие органические вещества на мономеры, которые всасываются плазмодием[5]. Разветвлённая эктоплазматическая сеть увеличивает поверхность всасывания[18]. У некоторых видов было описано фаготрофное питание[18].
Известно несколько ДНК- и РНК-содержащих вирусов, поражающих лабиринтул[23].
Размножение и жизненный цикл
[править | править код]Жизненный цикл лабиринтул не до конца понятен, но в нём, несомненно, имеет место половой процесс. У некоторых видов были описаны мейоз[18] и гаметы (женские и мужские гаметы неотличимы друг от друга, т. e. имеет место изогамия)[24]. У наиболее изученных представителей описаны многоклеточные цисты, спорангии (сорусы), а также двужгутиковые зооспоры и апланоспоры. При образовании спор клетки инцистируются или поодиночке, или группами, окружаясь тонкими стенками и дополнительно общей стенкой, так что образуется сорус. При благоприятных условиях отдельные цисты или каждая клетка соруса прорастают амёбоидными клетками или зооспорами (гораздо реже); в обоих случаях они дают начало новым плазмодиям[25].
Зооспоры имеют важное значение при определении родственных связей лабиринтул, а их строение указывает на принадлежность лабиринтул к страменопилам. Зооспоры представлены разножгутиковыми клетками, причём на переднем жгутике располагаются трёхчленные мастигонемы. Рулевой жгутик зооспор подстилается пигментным пятном[20].
Размножение происходит за счёт деления «клеток» внутри плазмодия надвое или путём разделения плазмодиев[20]. Плазмодий растёт при продолжающихся клеточных делениях и формировании новых слизистых тяжей[26].
Экология
[править | править код]Лабиринтулы — чрезвычайно важный компонент морского микробного сообщества. Они обитают в основном в морских и эстуарных местообитаниях, реже в пресных водоёмах или засоленных почвах. В 2014 году был описан новый пресноводный вид лабиринтул — Diplophrys mutabilis[27]. Лабиринтул можно обнаружить на водорослях, остатках сосудистых растений, в органическом детрите, где они образуют слизистые скопления[2][21].
Лабиринтулы играют важную роль в морских пищевых цепях. Представители Thraustochytrida часто становятся пищей амёб, а содержащиеся в них омега-3 полиненасыщенные жирные кислоты способствуют росту и размножению ракообразных. Многие морские рыбы, неспособные синтезировать эти соединения, вынуждены получать их из водорослей, на которых обитают лабиринтулы. Кроме того, лабиринтулы снабжают этими соединениями зоопланктон[18].
Хотя и в меньшей степени, чем бактерии, лабиринтулы могут выступать в роли первичных колонизаторов неорганических субстратов, в том числе пластика, стекла, алюминия и стеклоткани, оказавшихся погружёнными в воду. Личинки морского жёлудя Balanus amphitrite[англ.] лучше оседают на поверхностях, покрытых Thraustochytrida, чем на голых. Поэтому, возможно, лабиринтулы играют роль в обеспечении оседания личинок морских беспозвоночных[18].
Многие лабиринтулы являются комменсалами, паразитами или эндобионтами растений. Они способны пронизывать клеточную стенку у наземных растений, грибов, диатомовых (Thalassonema), зелёных (Cladophora) и жёлто-зелёных (Vaucheria) водорослей. Так, Labyrinthula zosterae и L. macrocystis вызывают карликовость нескольких видов морских трав семейства зостеровых[20]. По мнению ряда учёных, лабиринтулы ответственны за массовую гибель зостеры, неоднократно наблюдавшуюся на Белом море и морях Атлантического океана. Известны виды из наземных местообитаний — например, паразиты корней пшеницы[28]. Интересно, что некоторые виды Thraustochytrida хорошо развиваются на пыльце сосны, помещённой в морскую воду (что позволяет, в частности, их выделить для дальнейших исследований)[29][18].
Некоторые лабиринтулы освоили в качестве субстрата ткани и пищеварительные тракты животных и обитают там как паразиты, комменсалы или мутуалисты. Например, некоторые Thraustochytrida лучше изолируются с использованием личинок рачка Artemia вместо пыльцы сосны. Aplanochytrium yorkensis вызывает жаберные инфекции у двустворчатого моллюска Crassostrea virginica[англ.]. Некоторые лабиринтулы обитают в клетках печени у крабов. Лабиринтулы были также изолированы из тканей рыб, однако у них, судя по всему, они не вызывали заболевания[18]. Некоторые лабиринтулы являются патогенами кораллов-горгонарий Gorgonia ventalina[англ.][30].
Эволюционная история
[править | править код]У лабиринтул параллельно с животными, растениями и грибами наблюдается процесс выхода из воды на сушу, поскольку эктоплазматическая сеть позволяет сохранять условия водной среды и вне её. По этой причине лабиринтулы представляют большой эволюционный интерес[28]. Имеется ряд доказательств, указывающих, что лабиринтулы были первыми эукариотическими осмогетеротрофами. По-видимому, ранние лабиринтулы были способны как к осмотрофному питанию, так и к фаготрофному, что подтверждается способностью к фаготрофии у ряда современных видов. Кроме того, как упоминалось выше, траустохитриды могут расти на пыльце сосны. Это происходит благодаря их способности разрушать входящий в состав пыльцевых зёрен спорополленин[англ.], хотя он очень стоек к микробному разрушению. Способность к разрушению спорополленина говорит в пользу очень долгой эволюционной истории этой группы протистов[18].
Андерсон и Кавалье-Смит полагают, что предки лабиринтул по своей морфологии были близки к современным траустохитриевым, так что последние в наибольшей степени отражают исходное для класса состояние. Как тип строения лабиринтулид, так и тип строения Diplophrys и сходных с ним родов (Sorodiplophrys, Amphifila) являются вторичными, возникшими из типа строения ранних лабиринтул в ходе эволюции (причём последний из этих типов возникал независимо как минимум дважды)[17].
Систематика
[править | править код]История систематики
[править | править код]Олайв вынес класс Labyrinthulea в отдельный подтип Labyrinthulina типа Mycetozoa[14]. В большинстве классификаций, предложенных в последней трети XX века и самом начале XXI века, лабиринтул выделяли в отдельный тип (отдел). При этом микологи пользовались в качестве имени класса названием Labyrinthulomycetes (предложено Й. А. фон Арксом в 1970 году[31], валидизировано М. У. Диком в 2001 году[32]), а качестве названия отдела — названием Labyrinthulomycota (предложено Р. Х. Уиттекером в 1969 году[33]); первоначально объём обоих таксонов ограничивался лишь лабиринтуловыми (в 2003 году российский миколог Е. А. Кузнецов предложил выделить лабиринтул даже в отдельное царство Mycomyxina[13][34][35])[1]. По этому же пути пошли и протозоологи: когда в 1980 году Международный комитет по систематике простейших высказался за разделение ранее единого типа Protozoa на 7 типов, то класс Labyrinthulea был вынесен в отдельный тип Labyrinthomorpha Page, 1980[36][37].
С 1990-х годов, однако, всё большее распространение получают системы, в которых лабиринтулы рассматриваются именно в ранге класса, входящего в группу гетероконтов[38].
По данным филогенетического анализа 2009 года, лабиринтулы — монофилетическая группа. Данный анализ показал, что Labyrinthulidae и Aplanochytridae — сестринские группы, а Thraustochytrida (в традиционно принимавшемся объёме) — в действительности парафилетический таксон и распадается на две группы (Thraustochytrida1 и Thraustochytrida2)[13]. Thraustochytrida2 (группа, позднее выделенная в семейство Oblongichytriidae[17]) — сестринский таксон по отношению к группировке Labyrinthulidae и Aplanochytridae, а Thraustochytrida1 (остальные траустохитриды) — по отношению к общему предку Thraustochytrida2, Labyrinthulidae и Aplanochytridae[7].
Классификация 2012 года
[править | править код]По Андерсону и Кавалье-Смиту (2012), система класса Labyrinthulea выглядит следующим образом[17]:
Класс Labyrinthulea Olive, 1975 (лабиринтулы)
- Отряд Labyrinthulida Lankester, 1877 (лабиринтулиды)
- Семейство Labyrinthulidae Haeckel, 1868 (лабиринтуловые): род Labyrinthula[англ.]
- Семейство Aplanochytriidae Leander, 2001 (апланохитриевые): род Aplanochytrium
- Отряд Thraustochytrida Sparrow, 1973 (траустохитриды)
- Семейство Oblongichytriidae Cavalier-Smith, 2012 (облонгихитриевые): род Oblongichytrium
- Семейство Diplophryidae Cavalier-Smith, 2012 (диплофризовые): род Diplophrys
- Семейство Sorodiplophryidae Cavalier-Smith, 2012 (сородиплофризовые): род Sorodiplophrys
- Семейство Amphifilidae Cavalier-Smith, 2012 (амфифиловые): род Amphifila
- Семейство Althorniidae Cavalier-Smith, 2012 (алторниевые): род Althornia
- Семейство Thraustochytriidae Sparrow, 1943 (траустохитриевые): роды Thraustochytrium, Ulkenia, Schizochytrium[англ.], Japonochytrium, Aurantiochytrium, Sicyoidochytrium, Parietichytrium, Botryochytrium
Авторы системы преднамеренно трактуют отряд Thraustochytrida как парафилетический таксон (стремясь к большей однородности каждого из двух отрядов в плане морфологии)[17]. Альтернативой служит перенос семейства Oblongichytriidae в отряд Labyrinthulida, что делает оба отряда монофилетическими[39].
Дальнейшее развитие классификации
[править | править код]В 2013 году появились доводы в пользу включения в состав класса Labyrinthulea — в качестве наиболее рано отделившейся клады — вновь выделенной группы Amphitremida (амёбообразные простейшие из родов Amphitrema[англ.] и Archerella), представителей которой ранее относили к Cercozoa[39].
Предложенная Андерсоном и Кавалье-Смитом система класса Labyrinthulea с отрядами Labyrinthulida и Thraustochytrida была принята и в макросистеме живущих организмов, предложенной в 2015 году Майклом Руджеро с соавторами. Здесь данный класс рассматривается как один из шести классов типа Bigyra в надтипе Heterokonta[40][41]. При этом статус типа Bigyra остаётся пока неясным: по данным филогенетического анализа гетероконтов, результаты которого были опубликованы в 2013 году, это — парафилетический таксон, и класс Labyrinthulea вместе с плохо изученной и фенотипически неохарактеризованной группой Eogyrea образует в нём подтип Sagenista[англ.] (наиболее рано отделившуюся кладу гетероконтов), а остальные включаемые в тип Bigyra классы объединяются в надтип Opalozoa — сестринскую группу для клады, включающей два других типа гетероконтов (Pseudofungi и Ochrophyta); авторы анализа, впрочем, оговариваются, что имеются данные и в пользу монофилии типа Bigyra[42].
Классификация 2019 года
[править | править код]Группа систематиков в 2019 году предложила своё видение таксона и его связей[43]:39—40:
- Bigyra Cavalier-Smith, 1998 emend. 2006
- Opalozoa Cavalier Smith, 1991 emend. 2006
- Sagenista Cavalier-Smith, 1995
- Pseudophyllomitidae Shiratori et al., 2016
- Labyrinthulomycetes Dick, 2001
- Amphitremida Poche, 1913 emend. Gomaa et al., 2003: роды Amphitrema, Archerella, Diplophrys, Paramphitrema
- Amphifilida Cavalier-Smith, 2012: роды Amphifila, Fibrophrys, Sorodiplophrys
- Oblongichytrida Bennett et al., 2017: род Oblongichytrium
- Labyrinthulida Doffein, 1901: роды Aplanochytrium, Labyrinthula, Stellarchytrium
- Thraustochytrida Sparrow, 1943: роды Althornia, Aurantiochytrium, Botryochytrium, Japanochytrium, Monorhizochytrium, Parietichytrium, Schizochytrium, Sicyoidochytrium Thraustochytrium, Ulkenia
Хозяйственное значение
[править | править код]В настоящий момент лабиринтулы приобретают важное промышленное значение как природные источники омега-3-полиненасыщенных жирных кислот. Предлагается использовать лабиринтул для синтеза каротиноидов и сквалена, а также в составе кормов для аквакультуры[5]. Однако лабиринтулы могут вызывать заболевания у хозяйственно важных моллюсков, выращиваемых в марикультуре[18].
Примечания
[править | править код]- ↑ 1 2 3 4 5 Кусакин О. Г., Дроздов А. Л. Филема органического мира. Ч. 2. — СПб.: Наука, 1997. — 381 с. — ISBN 5-02-026018. — С. 240—246.
- ↑ 1 2 3 Хаусман и др., 2010, с. 109.
- ↑ Adl S. M., Simpson A. G., Lane C. E., Lukeš J., Bass D., Bowser S. S., Brown M. W., Burki F., Dunthorn M., Hampl V., Heiss A., Hoppenrath M., Lara E., Le Gall L., Lynn D. H., McManus H., Mitchell E. A., Mozley-Stanridge S. E., Parfrey L. W., Pawlowski J., Rueckert S., Shadwick R. S., Schoch C. L., Smirnov A., Spiegel F. W. The revised classification of eukaryotes // The Journal of Eukaryotic Microbiology. — 2012. — Vol. 59, no. 5. — P. 429—493. — doi:10.1111/j.1550-7408.2012.00644.x. — PMID 23020233.
- ↑ 1 2 Хаусман и др., 2010, с. 109—110.
- ↑ 1 2 3 4 Tsui C. K. M., Vrijmoed L. L. P. . A Re-Visit to the Evolution and Ecophysiology of the Labyrinthulomycetes // Marine Ecosystems / Ed. by A. Cruzado. — Winchester: InTech, 2012. — 310 p. — ISBN 978-953-51-0176-5. — P. 161—176.
- ↑ 1 2 Белякова и др., 2006, с. 46.
- ↑ 1 2 Tsui C. K. M., Marshall W., Yokoyama R., Honda D., Lippmeier J. C., Craven K. D., Peterson P. D., Berbee M. L. Labyrinthulomycetes phylogeny and its implications for the evolutionary loss of chloroplasts and gain of ectoplasmic gliding // Molecular Phylogenetics and Evolution. — 2009. — Vol. 50, no. 1. — P. 129—140. — doi:10.1016/j.ympev.2008.09.027. — PMID 18977305.
- ↑ Cienkowski L. Über den Bau und die Entwicklung der Labyrinthuleen // Archiv für mikroscopische Anatomie. — 1867. — Bd. 3. — S. 274–310.
- ↑ Olive, 1975, p. 215, 225.
- ↑ Sparrow F. K. Biological Observations on the Marine Fungi of Woods Hole Waters // The Biological Bulletin of the Marine Biological Laboratory, Woods Hole. — 1936. — Vol. 70, no. 2. — P. 236–263.
- ↑ Курс низших растений / Под ред. М. В. Горленко. — М.: Высшая школа, 1981. — 504 с. — С. 306—308.
- ↑ Olive, 1975, p. 228—230.
- ↑ 1 2 3 4 5 Белякова и др., 2006, с. 47.
- ↑ 1 2 Olive, 1975, p. 4—5, 215.
- ↑ Olive L. S. . Labyrinthulata // Synopsis and Classification of Living Organisms. Vol. 1 / Ed. by S. P. Parker. — New York: McGraw-Hill, 1982. — xlvii + 1119 p. — ISBN 0-07-079031-0. — P. 569.
- ↑ Cavalier-Smith T. The kingdom Chromista: origin and systematics // Progress in Phycological Research. Vol. 4 / Ed. by F. E. Round and D. J. Chapman. — Bristol: Biopress Ltd., 1986. — vii + 481 p. — ISBN 0-948737-03-4. — P. 309—347.
- ↑ 1 2 3 4 5 6 Anderson O. R., Cavalier-Smith T. Ultrastructure of Diplophrys parva, a new small freshwater species, and a revised analysis of Labyrinthulea (Heterokonta) (англ.) // Acta Protozoologica. — 2012. — Vol. 51, no. 4. — P. 291—304. Архивировано 4 марта 2016 года. — doi:10.4467/16890027AP.12.023.0783.
- ↑ 1 2 3 4 5 6 7 8 9 10 11 12 13 Seshagiri, Raghukumar. Ecology of the marine protists, the Labyrinthulomycetes (Thraustochytrids and Labyrinthulids) // European Journal of Protistology. — 2002. — Vol. 38, no. 2. — P. 127–145. — doi:10.1078/0932-4739-00832. Архивировано 24 сентября 2015 года.
- ↑ Белякова и др., 2006, с. 47—50.
- ↑ 1 2 3 4 Хаусман и др., 2010, с. 110.
- ↑ 1 2 3 Margulis, Chapman, 2009, p. 178.
- ↑ Белякова и др., 2006, с. 46—47.
- ↑ Takao Y., Tomaru Y., Nagasaki K., Honda D. Ecological Dynamics of Two Distinct Viruses Infecting Marine Eukaryotic Decomposer Thraustochytrids (Labyrinthulomycetes, Stramenopiles) // PLoS ONE. — 2015. — Vol. 10, no. 7. — P. e0133395. — doi:10.1371/journal.pone.0133395. — PMID 26203654.
- ↑ Margulis, Chapman, 2009, p. 181.
- ↑ Белякова и др., 2006, с. 47—49.
- ↑ Мюллер, Лёффлер, 1995, с. 169.
- ↑ Takahashi Y., Yoshida M., Inouye I., Watanabe M. M. Diplophrys mutabilis sp. nov., a new member of Labyrinthulomycetes from freshwater habitats // Protist. — 2014. — Vol. 165, no. 1. — P. 50—65. — doi:10.1016/j.protis.2013.10.001. — PMID 24334197.
- ↑ 1 2 Белякова и др., 2006, с. 48.
- ↑ Белякова и др., 2006, с. 50.
- ↑ Burge C. A., Douglas N., Conti-Jerpe I., Weil E., Roberts S., Friedman C. S., Harvell C. D. Friend or foe: the association of Labyrinthulomycetes with the Caribbean sea fan Gorgonia ventalina // Diseases of Aquatic Organisms. — 2012. — Vol. 101, no. 1. — P. 1—12. — doi:10.3354/dao02487. Архивировано 24 сентября 2015 года.
- ↑ Arx J. A. The Genera of Fungi Sporulating in Pure Culture. — Lehre: J. Cramer, 1970. — 288 p.
- ↑ Dick M. W. Straminipilous Fungi. Systematics of the Peronosporomycetes including Accounts of the Marine Straminipilous Protists, the Plasmodiophorids and Similar Organisms. — Dordrecht: Springer, 2001. — xv + 670 p. — ISBN 978-90-481-5639-9.
- ↑ Whittaker R. H. New Concepts of Kingdoms of Organisms // Science. — 1969. — Vol. 163, no. 3863. — P. 150–160. Архивировано 17 ноября 2017 года.
- ↑ Feofilova E. P. First Congress of Russian Mycologists // Microbiology. — 2002. — Vol. 71, no. 5. — P. 617–619. Архивировано 5 июня 2018 года.
- ↑ Кузнецов Е. А. Грибы водных экосистем // Диссертация на соискание учёной степени доктора биологических наук. — 2003. Архивировано 23 сентября 2015 года.
- ↑ Серавин Л. Н. Простейшие… Что это такое? / Отв. ред. Ю. И. Полянский. — Л.: Наука, 1984. — 176 с. — С. 5, 10, 188.
- ↑ Хаусман К. Протозоология. — М.: Мир, 1988. — 336 с. — ISBN 5-03-000705-9. — С. 36—37, 85.
- ↑ Хаусман и др., 2010, с. 96, 109.
- ↑ 1 2 Gomaa F., Mitchell E. A., Lara E. Amphitremida (Poche, 1913) is a new major, ubiquitous labyrinthulomycete clade // PLoS ONE. — 2013. — Vol. 8, no. 1. — P. e53046. — doi:10.1371/journal.pone.0053046. — PMID 23341921.
- ↑ doi:10.1371/journal.pone.09248
Вы можете подставить цитату вручную или с помощью бота. - ↑ Ruggiero M. A., Gordon D. P., Orrell T. M., Bailly N., Bourgoin T., Brusca R. C., Cavalier-Smith T., Guiry M. D., Kirk P. M. Correction: A Higher Level Classification of All Living Organisms (англ.) // PLOS One : journal. — 2015. — Vol. 10, iss. 6. — P. e0130114. — ISSN 1932-6203. — doi:10.1371/journal.pone.0130114. — PMID 26068874. — PMC 5159126.
- ↑ Cavalier-Smith T., Scoble J. M. Phylogeny of Heterokonta: Incisomonas marina, a uniciliate gliding opalozoan related to Solenicola (Nanomonadea), and evidence that Actinophryida evolved from raphidophytes // European Journal of Protistology. — 2013. — Vol. 49, no. 3. — P. 328—353. — doi:10.1016/j.ejop.2012.09.002. — PMID 23219323.
- ↑ Adl S. M., Bass D., Lane C. E., et al. Revisions to the Classification, Nomenclature, and Diversity of Eukaryotes (англ.) // Journal of Eukaryotic Microbiology : journal. — 2019. — Vol. 66, iss. 1. — P. 4—119. — ISSN 1066-5234. — doi:10.1111/jeu.12691. — PMID 30257078. — PMC 6492006.
Литература
[править | править код]Книги
[править | править код]- Белякова Г. А., Дьяков Ю. Т., Тарасов К. Л. . Ботаника: в 4 тт. Т. 2. Водоросли и грибы. — М.: Издат. центр «Академия», 2006. — 320 с. — ISBN 5-7695-2750-1. — С. 46—50.
- Хаусман К., Хюльсман Н., Радек Р. . Протистология / Пер. с англ. С. А. Карпова. Под ред. С. А. Корсуна. — М.: Товарищество научных изданий КМК, 2010. — 495 с. — ISBN 978-5-87317-662-5.
- Мюллер Э., Лёффлер В. . Микология. — М.: Мир, 1995. — 343 с. — ISBN 5-03-002999-0.
- Мухин В. А., Третьякова А. С. . Биологическое разнообразие: водоросли и грибы. — Ростов н/Д.: Феникс, 2013. — 269 с. — ISBN 978-5-222-20177-0.
- Margulis L., Chapman M. J. . Kingdoms & Domains: An Illustrated Guide to the Phyla of Life on Earth. 4th edition. — New York: W. H. Freeman and Company, 2009. — lxxii + 659 p. — ISBN 978-0-12-373621-5.
- Olive L. S. . The Mycetozoans. — New York: Academic Press, 1975. — x + 293 p. — ISBN 0-12-526250-7.
Статьи
[править | править код]- Ranasinghe C. P., Harding R., Hargreaves M. An improved protocol for the isolation of total genomic DNA from Labyrinthulomycetes // Biotechnology letters. — 2015. — Vol. 37, no. 3. — P. 685—690. — doi:10.1007/s10529-014-1712-1. — PMID 25355522.
- Gomaa F., Mitchell E. A., Lara E. Amphitremida (Poche, 1913) is a new major, ubiquitous labyrinthulomycete clade // PLoS ONE. — 2013. — Vol. 8, no. 1. — P. e53046. — doi:10.1371/journal.pone.0053046. — PMID 23341921.
- Ueda M., Nomura Y., Doi K., Nakajima M., Honda D. Seasonal dynamics of culturable thraustochytrids in estuarine and coastal waters (Labyrinthulomycetes, stramenopiles) // Aquatic Microbial Ecology. — 2015. — Vol. 74, no. 3. — P. 187—204. — ISSN 0948-3055. — doi:10.3354/ame01736. Supplementary material 1.
Ссылки
[править | править код]- Класс Labyrinthulea (англ.) в Мировом реестре морских видов (World Register of Marine Species). (Дата обращения: 18 февраля 2020).
Классификация Sar | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Страменопилы |
| ||||||||||||||||
| Альвеоляты |
| ||||||||||||||||
| Ризарии |
| ||||||||||||||||
Эта статья входит в число хороших статей русскоязычного раздела Википедии. |