French
French Deutsch
DeutschТънко черво – Уикипедия
| Тънко черво | |
|---|---|
 | |
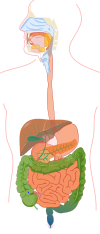
| Схематично изображение на тънките черва | |
| Латински | intestinum tenue |
| Атлас | Тънко черво |
| Артерия | Arteria mesenterica superior |
| Вена | Vena portae |
| Нерв | Ganglia celiaca, Nervus vagus[1] |
| Лимфа | Truncus intestinalis |
| MeSH | Small+intestine |
| Elsevier | Small intestine |
| Тънко черво в Общомедия | |

празно черво (jejunum)
и хълбочно черво (ileum)
Тънкото черво (на латински: intestinum tenue) е част от храносмилателния канал на гръбначните животни включително и на човека. Представлява дълга нагъната тръба, която започва от пилора на стомаха и завършва при илео-цекалната клапа, където е границата му с дебелото черво. В тънкото черво се извършва основно храносмилане на непреработените в стомаха вещества и адсорбцията им до по-нискомолекулни вещества и минерални соли.[2] В началните отдели на тънкото черво се намират изводните канали на две от най-големите жлези в организма – черен дроб и панкреас, както и се отделя секрет от множество микроскопични жлези разположени в подлигавичния слой на органа.
Анатомично устройство
[редактиране | редактиране на кода]Тънкото черво се разполага в централната част на коремната кухина, силно нагънато. Относителната дължина при зрял мъж е 6,9 m, а при жената е 7,1 m. Този размер обаче може да варира в големи граници от 4,6 до 9,8 m.[3][4] Средният диаметър на просвета е 2,5 – 3 cm, като най-голям е в областта на дванадесетопръстника, около 4 – 5 cm, като към края си постепенно намалява и достига около 3 cm.
Тънкото черво има три части:
- Дванадесетопръстник (на латински: duodenum) – обхваща първите 25 – 30 cm от тънкото черво. Тук се отваря жлъчният канал и каналът на задстомашната жлеза.
- Празно черво (на латински: jejunum) – с дължина 2/5 от останалата част.
- Хълбочно черво (на латински: ileum) – с дължина 3/5 от останалата част.
Празното черво и хълбочното черво са разположени интраперитонеално и са свързани с коремницата чрез нейна дубликатура, наречена опорак или мезентериум (на латински: mesenterium). По своето протежение те образуват множество нагъвания, наречени бримки. Бримките са разположени повърхностно (около 1/3 от тях) и дълбоко (останалите 2/3), като бримките на празното черво имат предимно хоризонтално направление, а на хълбочното предимно вертикално. Празното черво с преимущество се разполага предимно наляво, а хълбочното предимно надясно от срединната линия.
Тънкото черво влиза в пространствени отношения спрямо останалите органи в коремната кухина. Отдясно и отляво то е заобиколено съответно от възходящата и низходящата част на дебелото черво. Отпред е покрито от голямото було. Назад то влиза в досег с дванадесетопръстника, главата на задстомашната жлеза, черния дроб, бъбреците, коремната аорта, долната куха вена и т.н.
Надолу, в областта на малкия таз, то е в досег с пикочния мехур и матката с нейните структури при жените.
Хистологичен строеж
[редактиране | редактиране на кода]
В лигавицата на тънкото черво се намират много жлези, които отделят чревен сок. Именно в нея става всмукването на продукти от разграждането на белтъците, мазнините и въглехидратите. Тя е образувана от малки власинки, достигащи на височина до 1 mm, и увеличаващи вътрешната повърхност на тънкото черво около 6 пъти, което е от голямо значение за всмукването. Във всяка една власинка има кръвоносни и лимфни съдове, както и гладки мускулни влакна. Власинките могат да се свиват и отпускат, както и да извършват движение в страни, като по този начин улесняват всмукването на хранителни и други вещества. През клетките от епитела, образуващ стената на власинките, преминават не всички, а само някои хранителни вещества. Това преминаване става винаги в една посока – от червото към власинките.
Трите части на тънките черва имат сходен строеж и са подобни на външен вид при разглеждане под микроскоп, но има известни различия, резюмирани в таблицата:
| Слой | Дванадесетопръстник | Празно черво | Хълбочно черво |
| Серозна мембрана | нормална | нормална | нормална |
| Външен мускулен слой | надлъжни и пръстеновидни слоеве, с Ауербахови нервни сплитове между слоевете | аналогично на дванадесетопръстника | аналогично на дванадесетопръстника |
| Субмукоза | Брунерови жлези и Мейснерови нервни сплитове | без Брунерови жлези | без Брунерови жлези |
| Мукоза: Мускулна мукоза | нормална | нормална | нормална |
| Мукоза: Ламина проприя | без Пайерови плаки | без Пайерови плаки | Пайерови плаки |
| Мукоза: Чревен епител | прост чревен епител, съдържащ клетки на Пенет, както и бухалковидни клетки | подобно на дванадесетопръстника; чревните вили са дълги | подобно на дванадесетопръстника; чревните вили са къси |
Ако червата се разгледат като цилиндър, дълъг 280 cm, с диаметър 4 cm, повърхностната площ би била 3300 cm². Гънките увеличават повърхността три пъти, което прави площ от 10 000 cm². Власинките допълнително увеличават площта десетократно.
Физиологични особености
[редактиране | редактиране на кода]Храната постъпва в тънките черва през пилорния отвор на стомаха при отварянето на пилорния сфинктер, след което с посредством вълнообразни движения се придвижва надолу по дължината му към дебелото черво, чрез т. нар. чревна перисталтика. Тези движения се регулират от нервната система, а механичното дразнение на хранителния химус върху лигавицата предизвиква по рефлексен път усилване на перисталтичните движения. Зеленчуците и плодовете предизвикват по-силни дразнения на червото и съответно по-силна перисталтика. Също така различни химични вещества могат по кръвен път да регулират движенията на тънкото черво. При приемане на развалена храна в стомаха, както и при претоварване с повече храна, перисталтиката на тънкото черво може да се усили и да се появи диария.
С малки изключения по-голямата част от поетата храна при животните поетата храна се разгражда в чревния канал до нейните основни съставки. Получените крайни продукти се резорбират и навлизат в кръвта и лимфата. Навлизайки през пилора към дванадесетопръстника хранителното съдържание има вид на каша, наречена химус. Тук той е подлаган на въздействието на панкреатичния сок, жлъчката и чревния сок под влияние на които сложните хранителни вещества се хидролизират до техните основни продукти. Само част от поетата храна достига до дебелите черва, където се подлага на разграждане. Освен смилателна функция тънките черва имат и резорбтивна функция. За ефективното съчетаване на двете потокът на съдържанието трябва да се регулира така, че да осигурява смесване на луменовото съдържание с ензимите на чревния сок и смилането на мазнините, въглехидратите и белтъчините, както и максималната им експозиция върху лигавицата на чревния лумен. Скоростта на изпразване на тънките черва е от първостепенна важност за предотвратяване на тяхното препълване. Преминаването през илеума се забавя в резултат на по-високия процент контракции на сегментите в илеума в сравнение с тези в иеюнома. Илеоцекалната клапа също до известна степен забавя преминаването.
Тънкото черво извършва три вида движения, които са резултат на взаимното координиране на надлъжната и циркулярната мускулатура:
- Махаловидни движения. Те наподобяват движението на махало и са също ритмични, периодични съкращения на по-големи участъци. При тях се съгласува двигателната дейност и на двата слоя чревни мускули. Надлъжните мускули ритмично се свиват и се отпускат, като на известно протежение тънкото черво ту се скъсява, ту се удължава и неговото съдържимо се премества ту напред, ту назад. Отделните частици на чревното съдържимо не се придвижват по дължината на червата, а правят движения, подобни на махало, откъдето произлиза и името махаловидни движения. Благодарение на тези движения съдържимото се смесва по-добре с чревния сок, постига се размесване на чревното съдържимо и се улеснява резорбцията на хранителните вещества.
- Перисталтични движения. Представлява последователното съкращение на пръстеновидния слой по протежение на червата от оралния към аналния край, като съседният, дистално разположен участък се отпуска, осигурява еднопосочното придвижване на хранителния химус. Перисталтичната активност се усилва значително след хранене. Това се дължи отчасти на постъпването на химус в дванадесетопръстника, а също и на разтягането на стомаха, така че се включва и гастро-ентерален рефлекс.
- Сегментни движения. Представляват ритмични посменни свивания и разпускания на пръстеновидните мускули в близки участъци. Тези движения са предимно в резултат на периодичното свиване на пръстеновидната мускулатура, което става в различни участъци на сегмента. По този начин чревното съдържимо се размесва с храносмилателните сокове и настъпва известно забавяне на преминаването на съдържанието по време на смилането.
Основната регулаторна система за движението на тънките черва е електрическата бавна вълна. Първичният пейсмейкър е разположен в надлъжната мускулатура на дванадесетопръстника близо до входа на жлъчния канал. Бавните вълни се генерират от пейсмейкъра с честота например 17 – 18 за минута при кучето и котката и 14 – 15 при коня. Тези вълни се придвижват по цялата дължина, но локални пейсмейкъри в най-отдалечените области произвеждат свои бавни вълни с редуцирана честота. Така към задните отдели на тънкото черво бавните вълни намаляват по честота и скорост на разпространение. Така например в илеума на коня те стават 10 – 11 в минута. Движенията се регулират както от вътрешна, така и от външна инервация, която може да увеличи или намали вероятността от залпово изпразване на червата. Най-важният вътрешен рефлекс е перисталтичният. Перисталтиката се дължи на последователни контракции на надлъжната и пръстеновидната мускулатура и индуцира масово преминаване на чревното съдържание. Свиването на надлъжната мускулатура и потискането на пръстеновидния мускул предизвиква скъсяване на червото и разширява лумена му. Същевременно настъпва отпускане на надлъжната мускулатура и свиване на пръстеновидната. Това намалява лумена и придвижването на храната назад. Възбудители на движенията на тънките черва са преди всичко местните механични и химична дразнения, т.е. плътността на хранителната кашица и нейният химичен състав. Колкото по-голямо е количеството на плътните, неразтворими във вода или несмилаеми частици в чревното съдържимо, толкова по-усилена е чревната перисталтика. Оттук и значението на „баластните хранителни вещества“ и на първо място на целулозата, които, увеличавайки плътността на чревния химус, значително усилват перисталтиката.
Съществуват и външни инхибиторни рефлекси, причинени от болка или подуване и могат да потиснат движенията. Рефлексите се провеждат от вагални аферентни нерви към блуждаещия нерв, които провеждат стимула от механорецепторите до нервния център като вагалната центробежна стимулация е предимно възбуждаща.
Ендокринната регулация на двигателната активност на червата се стимулира от хормоните гастрин и холецистокинин, а секретинът потиска контракциите на мускулатурата.
Храносмилане
[редактиране | редактиране на кода]Тънките черва са мястото, където протича по-голямата част от химическото разграждане на хранителните вещества. Повече храносмилателни ензими се секретират от панкреаса и преминавайки през панкреатичния канал се изливат в началните части на тънкото черво. Ензимите се изливат в отговор на хормона холецистокинин, произвеждан от тънките черва, като резултат от наличието на храна. Друг хормон, наречен секретин, е причината за секрецията на бикарбонати с цел неутрализация на киселините постъпващи от стомаха. Трите основни класа биохимични съединения, които търпят разграждане в тънките черва са белтъците, липидите и въглехидратите:
Смилане на белтъците
[редактиране | редактиране на кода]Белтъците и пептидите се разграждат до аминокиселини. Химическата хидролиза започва от стомаха и продължава в проксималната част на тънките черва. Протеолитичните ензими на тънките черва, като например трипсин и химотрипсин, секретирани от панкреаса са причина за раздробяването на белтъците от храната до малки пептидни фрагменти. Други важни ензими са карбоксипептидазата, освобождаваща свободни аминокиселини, както и ензимите аминопептидаза и дипептидаза, хидролизиращи пептидните фрагменти до свободни аминокиселини.
Резорбция на аминокиселини се осъществява навътре в епителните клетки чрез специални транспортни механизми по начин подобен на хексозния транспорт. Има поне две отделни системи на активен транспорт – една за неутралните и една за основните аминокиселини. Установени са и системи за транспортиране на дипептиди и трипептиди в клетката. Над 85% от хидролазната активност спрямо дипептидите е локализирана в цитоплазмата на клетките, така че около 90% от пептидите, които влизат в нея се хидлоризират по-нататък до свободни аминокиселини. Останалите 10% нехидролизирани пептиди могат да дифундират през базолатералната мембрана в кръвта.
Смилане на липидите
[редактиране | редактиране на кода]Липидите се разграждат до мастни киселини и глицерол. Панкреатичната липаза е ензимът на тънките черва, който хидролизира триглицеридите до свободни мастни киселини и моногрицериди. Този ензим функционира с помощта на жлъчните сокове, секретирани от черния дроб и жлъчния мехур. Жлъчните соли емулгират триглицеридите, като ги правят достъпни за панкреатичната липаза. Това е необходимо, тъй като липазата е водно-разтворима, докато мастните киселини на триглицеридите са хидрофобни и съответно неразтворими във вода. Жлъчните соли са помощник, доближаващ двете фази – триглицеридите и ензима панкреатична липаза. За процеса на смилане на мазнините се изисква повече време, отколкото за въглехидратите и протеините. При превръщането на мазнините от емулсия в мицел диаметърът на частиците намалява 100 пъти, а повърхността се увеличава над 10 000 пъти. Мицелът дифундира от емулсионните частици във власинковия слой на епитела, където мазнината се отделя, за да дифундира през липидната мембрана в клетката.
Смилане на въглехидратите
[редактиране | редактиране на кода]Някои въглехидрати се разграждат до прости монозахариди. Панкреатичната амилаза е отговорна за разграждането на въглехидратите (например, скорбяла) до олигозахариди. Други въглехидрати остават неразградени при преминаването си през тънките черва, но тяхното разграждане става в дебелото черво с помощта на бактерии. Някои въглехидрати, като например целулозата, не се разграждат и усвояват, въпреки че са изградени от глюкозни мономери.
При смилането на въглехидратите в тънките черва има две общи фази – луменна и мукозна. Въглехидратите в храната са главно нишестета, захароза, лактоза и нишковидни въглехидрати. Продуктите от луменната фаза на смилане не могат да се резорбират от мукозата, а трябва да се разградят по-нататък до монозахариди, които могат да се транспортират в епителната клетка. Мукозната фаза на това разграждане се извършва във власинковия слой на епитела. Ензимите са инкорпорирани в луменната мембрана на клетката и мигрират към връхчето на микровилите.
| Източник | Ензим | Активатор | Субстрат | Действие |
| Екзокринен панкреас | Трипсин | Ентерокиназа | Протеин | Разкъсва пептидните връзки, образувани от карбоксилните групи на аргинина и лизина. |
| Химотрипсин | Трипсин | Протеин | Разкъсва пептидните връзки, образувани от карбоксилните групи на аминокиселините. | |
| Еластаза | Трипсин | Протеин | Разкъсва пептидните връзки, образувани от карбоксилните групи. | |
| Карбоксипептизаза А | Трипсин | Протеин | Откъсва крайните аминокиселини. | |
| Карбоксипептизаза Б | Трипсин | Протеин | Откъсва крайните аминокиселини. | |
| Панкреасна липаза | Триглицериди | Разгражда моноглицеридите до мастни киселини. | ||
| Панкреасна естераза | Холестеролови естери | Разгражда холестерол до мастни киселини. | ||
| Фосфолипаза А2 | Трипсин | Лецитин | Лизолецитин | |
| Панкреасна α-амилаза | Йони на хлора | Нишесте | Хидролизира α-1,4 връзките, като образува α-лимит декстрини, малтотриоза и малтоза | |
| Рибонуклеаза | РНК | Нуклеотиди и олигонуклеотиди. | ||
| Дезоксирибонуклеаза | ДНК | Нуклеотиди и олигонуклеотиди. | ||
| Чревна мукоза | Ентеропептидаза | Трипсиноген | Трипсин | |
| Аминопептидаза | Пептиди | Откъсва N-крайните аминокиселини от пептидите. | ||
| Дипептидази | Дипептиди | Две аминокиселини. | ||
| Малтаза | Малтоза | Глюкоза. | ||
| Лактаза | Лактоза | Галактоза и глюкоза. | ||
| Захараза | Захароза | Фруктоза и глюкоза. | ||
| Фосфолипаза | Фосфолипиди | Глицерол, мастни киселини, фосфати. | ||
| Нуклеаза, нуклеотидаза и нуклеозидази | Нуклеинова киселина, нуклеотиди, нуклеозиди | Холи или други основи. |
Абсорбция
[редактиране | редактиране на кода]Разградените хранителни вещества са способни да навлязат през стените на кръвоносните съдове чрез процес на дифузия. Тънките черва са мястото, където по-голяма част от хранителните вещества се абсорбират от организма. Стената на тънките черва (чревната мукоза) е разграничена от празнината на тънкото черво, посредством слой от клетки на цилиндричния епител. Чревната мукоза е нагъната, което увеличава абсорбционната повърхност. Повърхността на гънките е покрита с микроскопични власинки, наречени чревни вили, а отделните епителни клетки на вилите също имат власинковидни образования, наречени микровили. Функциите на гънките, вилите и микровилите е да увеличат до максимум контактната повърхност с хранителните вещества.
В сърцевината на всяка вила има система от капиляри и тънки лимфни съдове. Епителните клетки на вилите поемат и транспортират хранителните вещества от празнината на тънкото черво до капилярите в сърцевината на вилата. Абсорбираните вещества се транспортират от капилярите по кръвоносната система до други части на тялото, където се използват за строежа но нови биомолекули. Храната, която остава неразградена и неусвоена, навлиза към дебелото черво.
Абсорбцията на повечето хранителни вещества се извършва в празното черво (jejunum), с изключение на желязото което се абсорбира в дванадесетопръстника, витамин B12 и жлъчните соли, абсорбирани в крайните части на хълбочното черво.
Водата и липидите се абсорбират посредством пасивна дифузия, натрия се абсорбира посредством активен транспорт, глюкозата и аминокиселините чрез котранспорт, а фруктозата – чрез улеснена дифузия.
При други животни
[редактиране | редактиране на кода]Тънко черво се открива при всички четирикраки, както и при висшите костни риби (Teleostei), въпреки че формата и размерите варират при различните видове. При костните риби е сравнително късо, обикновено под два пъти дължината на тялото и има редица гънки (caeca pylori), които увеличават общата повърхност. При тези риби, липсва илео-цекална клапа на границата между тънкото и дебелото черво.[5]
При четирикраките винаги се открива наличие на илео-цекална клапа, отваряща се в дебелото черво. Дължината на тънките черва при четирикраките е значителна в сравнение с тази при костните риби, като по правило растителноядните животни, както и бозайниците и птиците (топлокръвни животни) имат по-дълги тънки черва отколкото земноводните и влечугите (студенокръвни животни). Контактната повърхност при всички четирикраките е увеличена в резултата на наличието на множество гънки, като при бозайниците има и същински чревни вили.[5]
Границите между дванадесетопръстника, празното и хълбочното черво са едва доловими при хората, а при останалите животни обикновено не се прави стриктно разграничение или се приемат условни граници.[5]
Останалите риби, като например акулообразните, есетроподобните и ръкоперките не притежават тънко черво. Вместо това при тях стомаха се свързва с ректума, посредством така нареченото спирално черво. При този тип чревна система, чревната празнина е относително права, но притежава гънки по дължина, спираловидно усукани около централната ос. По този начин също има увеличение на абсорбционната повърхност.[5]
При миногите, спирално разположените гънки са сравнително малко, тъй като храната на този тип риби изисква минимално храносмилане. Миксините не притежават спирални гънки, като разграждането на храната става по цялото протежение на чарвата, които не са обособени на различни части.[5]
| Част на червото | Говедо | Овца и коза | Свиня | Куче | Котка | Кон |
|---|---|---|---|---|---|---|
| Тънко черво | 27 – 49,2 | 18,1 – 35,2 | 16 – 21 | 1,8 – 4,8 | 0,8 – 1,3 | 19 – 30,3 |
| Дванадесетопръстник | 0,9 – 1,2 | 0,6 – 1,2 | 0,7 – 0,95 | 0,2 – 0,6 | 0,1 – 0,12 | 1,3 – 1,5 |
| Празно и хълбочно черво | 26,1 – 48 | 17,5 – 34 | 15,3 – 20 | 1,6 – 4,2 | 0,7 – 1,2 | 17 – 28 |
Тънкото черво при говедото заема около 79% от общата дължина на храносмилателния канал. При овцата и козата е около 81%, при домашната свиня – 79%, при коня – 76%, при кучето – 86%, при котката е около 75%.
Имунология на тънките черва
[редактиране | редактиране на кода]Храносмилателният тракт притежава голяма епителна повърхност, подложена на действието на антигенни структури на външната среда. Те биват от различно естество – антигени на храната, антигени на микробната микрофлора, на патогенните микроорганизми и на различни видове паразити. Поради тази причина в стомашно-чревната мукоза има голямо количество имунологично активни клетки – макрофаги, лимфоцити, еозинофили, неутрофили и мастоцити. Благодарение на тях чревната мукоза може да формира имунен отговор спрямо попадналите в нея антигени. Имунологично активните клетки са разположени в три основни структури на тънките черва – Пайерови плаки, дифузно в ламина проприя и епителиума на чревните власинки.
Пайеровите плаки представляват групи от лимфоидни фоликули. Разположени са в мукозата и субмукозата и подобно на сливиците са част от т.нар. организирана лимфоидна тъкан. Те играят роля на отключващ момент за имунния отговор на червата. Епителът на техните фоликули съдържа М-клетки, които са се специализирали в разкриване на антигените попаднали в чревния лумен. Под епитела им се намират и лимфоцитите и макрофагите. През него преминават интактни белтъчни молекули и вируси, които биват унищожавани от горепосочените клетки. Така се формира локалният имунен отговор. Лимфоидните фоликули на Пайеровите плаки съдържат голямо количество В- и Т-лимфоцити, които в по-голямата си част постъпват в кръвта. След съзряването си те произвеждат имуноглобулини, които определят хуморалната защита на организма. В-лимфоцитите преминават в ламина проприя на тънките черва, дихателните пътища, гениталния тракт и млечната жлеза при женските организми.
Хуморалният механизъм на имунния отговор в червата се осъществява от освободени молекули на имуноглобулин А (IgA). Те преминават в чревния лумен като се свързва с патогенни бактерии, токсини, протозойни паразити и други.
В следродилния период всички майчини имуноглобулини при преживните животни, прасетата и конете се получават от коластрата посредством чревния предавателен комплекс. Тези животни са хипогамаглобулинни при раждането си. По време на непосредствения постнатален период има къс интервал, през който чревната мукоза е високопроницаема за всички макромолекули в контакт с мукозата. Увеличеното приемане на имуноглобулини от тънките черва трае много кратко време като след това червата се „затварят“, за да предотвратят по-нататъшното масово приемане на протеини.
Някои от гризачите и насекомоядните бозайници развиват пасивен имунитет както чрез интраутеринен транспорт, така и чрез следродилна чревна резорбция. Тези видове имат удължен период на селективен транспорт на гамаглобулини.
При човека пасивен имунитет се придобива почти изцяло чрез интраутеринен транспорт и съществува слаба абсорбция на гамаглобулини през тънките черва
| Видове | Преднатално | Постнатално |
|---|---|---|
| Кон | 0 | *** (24 h) |
| Свиня | 0 | *** (24 – 36 h) |
| Говедо, овца, коза | 0 | *** (24 h) |
| Куока (на латински: Setonix brachyurus) | 0 | *** (180 дни) |
| Куче, котка | * | ** (1 – 2 дни) |
| Домашни птици | ** | ** (5 дни) |
| Таралеж | * | ** (40 дни) |
| Мишка | * | ** (16 дни) |
| Плъх | * | ** (20 дни) |
| Морско свинче | *** | 0 |
| Домашен заек | *** | 0 |
| Човек | *** | 0 |
Литература
[редактиране | редактиране на кода]- Гаджев С. „Приложна и топографска анатомия на домашните животни и птици“, Стара Загора, 1995, ISBN 954-8180-31-6
- Германов А. „Приложна зоология“. Земиздат, 1992, ISBN 954-05-0187-3
- Големански В., „Зоология Безгръбначни животни“, УИ „Епископ Константин Преславски“, второ издание 2003, ISBN 954-577-187-9
- Иванов В., „Патологична физиология на домашните животни“, Земиздат, София 1995 г.
- Ковачев Г., Гигов Ц., „Анатомия на домашните животни“, том I, Земиздат, София 1995 г. ISBN 954-05-0330-2
- Коларов Я., „Зоология“, УИ „Паисий Хилендарски“, 2008, ISBN 978-954-423-421-8
- Кръстев Х., Витанов С., „Ембриология“, Земиздат, София 1994 г. ISBN 954-05-0203-9
- Кръстев Х., Витанов С. „Цитология и Хистология“. Земиздат, София, 1993, ISBN 954-05-0083-4
- Томов Тр., Седлоев Н., Градинарски Г., Костов Й., Илиев Я., Биволарски Б., Георгиев П., „Ветеринарномедицинска физиология“, Издателство Тракийски университет, Стара Загора 1998 г.
Източници
[редактиране | редактиране на кода]- ↑ Medical College of Georgia 6/6ch2/s6ch2_30
- ↑ www.britannica.com
- ↑ Elsevier: Gray's Anatomy, 40th Edition
- ↑ Lea Brothers and Co. 1907: Surgical Applied Anatomy
- ↑ а б в г д Romer, Alfred Sherwood. The Vertebrate Body. Philadelphia, PA, Holt-Saunders International, 1977. ISBN 0-03-910284-X. с. 349 – 353.
| ||||||||||||||||||||||